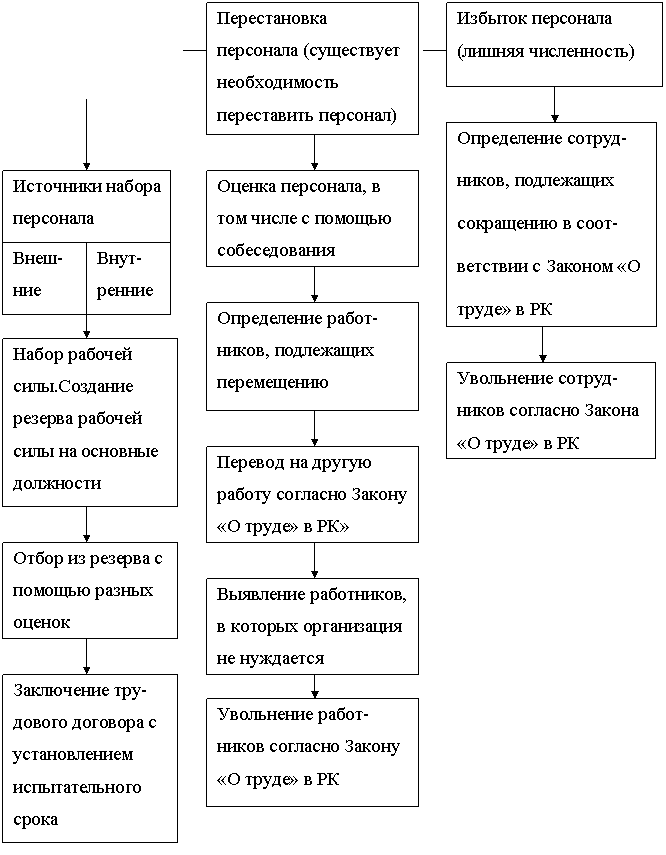
Перестановка кадров: 6. Ротация и перестановка кадров. Должностная инструкция руководителя, или «Управленческая восьмёрка»
Содержание
6. Ротация и перестановка кадров. Должностная инструкция руководителя, или «Управленческая восьмёрка»
Читайте также
Текучесть кадров
Текучесть кадров
В современном обществе «текучка» кадров – частая проблема. Она зависит от множества факторов, на многие из которых можно повлиять. На некоторые повлиять нельзя, но можно изменить отношение сотрудника к этим факторам.Факторы, влияющие на текучесть
16.7. Инверсия (перестановка)
16.7. Инверсия (перестановка)
Изменение привычного порядка слов для того, чтобы подчеркнуть какую-либо мысль. К примеру, сказуемое идет впереди подлежащего. Для тех, кто забыл эти термины школьной программы, скажем проще. Когда глагол идет перед существительным. Но это не
Сохранение кадров
Сохранение кадров
В компании Southwest – самый низкий коэффициент текучести кадров во всей отрасли, хотя у некоторых сотрудников оплата труда ниже, чем в других авиакомпаниях на тех же самых должностях. Это свидетельствует о том, что люди работают там, потому что им там
Это свидетельствует о том, что люди работают там, потому что им там
8.4.1. Ротация труда
8.4.1. Ротация труда
Работники фактически на всех уровнях организации могут быть ротированы через ряд рабочих мест с тем, чтобы расширить свои знания и навыки. Ротация труда обеспечивает организации большую гибкость, потому что обязанности работников могут быть легко
9.2.2. Ротация труда
9.2.2. Ротация труда
Ротация работы – это систематический перевод сотрудников с одной должности на другую, что позволяет увеличить число заданий, выполняемых одним человеком, без повышения сложности работы. Например, рабочий завода по сборке автомобилей в течение одной
Ротация труда
Ротация труда
Ротация труда (работы) – это систематический перевод сотрудников с одной должности на другую, что позволяет увеличить число заданий, выполняемых одним человеком, без повышения сложности работы. Например, рабочий завода по сборке автомобилей в течение
Например, рабочий завода по сборке автомобилей в течение
Ротация труда
Ротация труда
Работники фактически на всех уровнях организации могут пройти ротацию через ряд рабочих мест и расширить свои знания и навыки. Ротация труда обеспечивает организации большую гибкость, потому что обязанности работников могут быть легко изменены в связи с
5. Текучесть кадров
5. Текучесть кадров
В разных национальных культурах есть свои представления о том, сколь долго человек должен работать на одном месте, чтобы счесть его желанным кандидатом для своей компании.В России принято считать, что человек надежен, если он работает на одном месте
Независимость от кадров
Независимость от кадров
ПРИМЕР 1
Эта история случилась в региональном представительстве моего бизнеса в Тольятти. Перспективная сотрудница утром ехала на работу на своей машине. Доехала до очередного перекрестка, встала на красный свет. А по перпендикулярной дороге
Перспективная сотрудница утром ехала на работу на своей машине. Доехала до очередного перекрестка, встала на красный свет. А по перпендикулярной дороге
Ротация кадров
Ротация кадров
Для хорошей работы бизнеса нужна ротация кадров. Если мы не можем менять ключевых сотрудников, увольняем персонал, который не жалко. В том числе и в показательных целях.Некоторые новички, конечно, могут не выдержать такого темпа. У нас был случай, когда
Кузница кадров
Кузница кадров
Очень важно создать собственную кузницу кадров. Для этого введите «черновую» позицию для обучения – позицию стажера, на которую вы постоянно набираете людей. Например, есть люди, которые делают холодные звонки. Для выполнения этой работы вы можете брать
Организация обеспечила подготовку сотрудников и предоставила оборудование для укрепления базы четырех общинных радиостанций в [. Карибском бассейне («Roоts FM», Ямайка; «Radio […] Muye», Суринам). unesdoc.unesco.org | The Organization also provided training and equipment to reinforce the capacity of four community radio […] stations in the Caribbean (Roots FM, Jamaica; Radio Paiwomak, Guyana; […] Suriname). unesdoc.unesco.org |
| Добавить код BF к соответствующим номерам […] заказов муфт и ниппелей. staubli.com | Add the code BF to the concerned part-numbers […] of the sockets and the plugs. staubli.com |
RFLQ_S007BA Расчет ликвидности: [. ..] ..]перенести фактические данные в нов. бизнес-сферу . enjoyops.de enjoyops.de | RFLQ_S007BA Liquidity Calculation: […] Transfer Actual Data to New Business Area . enjoyops.de enjoyops.de |
| RM06BA00 Просмотр списка заявок . enjoyops.de enjoyops.de | RM06BA00 List Display of Purchase Requisitions . enjoyops.de enjoyops.de |
| Эта опция меню будет доступна после установки CD/DVD/BD—ROM-привода в NMT, или при подключении внешнего USB-привода CD/DVD/BD—ROM. popcornhour.es | This option will only be accessible when a CD/DVD/BD-ROM drive has been installed into or attached to your NMT. popcornhour.es |
Компания также поставляет систему шасси для первого в мире гражданского конвертоплана «Tiltrotor» […] вертикально взлетающего вертолёта. safran.ru | It also supplies the landing gear for the Bell/Agusta Aerospace BA609, the world’s first civilian tilt-rotor aircraft, combining the flexibility of vertical flight with the speed and range of a conventional aircraft. safran.ru |
Во-вторых, […] ветра шасси используется в основном для различных кондиционеры, воздушные [. завесы, отопления и охлаждения, вентилятор и т.д., также могут быть использованы в промышленных и горнодобывающих предприятий, общественных мест, крытый вентиляции. ru.shyngda.com | Second, use VAV BF type low-noise wind […] chassis is mainly used for a variety of air conditioning units, air curtain, heating […] and cooling fan, etc., can also be used in industrial and mining enterprises, public places, indoor ventilation. en.shyngda.com |
Чтобы привести автомобиль в боевую готовность и показать силу были использованы 3-дюймовые навесы и особые […] колеса матового черного цвета, а также […] большой передний кенгурятник, ограничительная […] планка и багажник на крыше. ms-auto.co.jp | To be fully armed and show the impact, 3 inch lift ups and [. special mat black wheel and BF Goodrich […] and tail guard and roof racks are added. ms-auto.co.jp |
Еще одним из популярных туристических мест в 2010 […] tourism-review.ru | Among other popular destinations for 2010 will be, […] tourism-review.com |
| BD выпускается в строгом соответствии с техническими условиями, все аудио могут быть расшифрованы вывода см. в разделе BD RIP, BD ISO треков были совершенны следующего поколения выходе источника macbook-covers. | BD produced in strict accordance with specifications, all the audio can be decoded output, see BD RIP, BD ISO tracks were perfect the next generation of source output macbook-covers.net |
Рейтинг финансовой устойчивости […] Ардшининвестбанку как одному из крупнейших […] банков Армении (будучи вторым банком в Армении по величине активов с долей рынка в 12,2% в 2007 году, Ардшининвестбанк в марте 2008 года стал лидером по этому показателю), широкой филиальной сетью, хорошими финансовыми показателями, особенно – растущей рентабельностью, высокой капитализацией и показателями эффективности выше среднего в контексте армянского рынка. ashib.am | According to Moody’s, ASHIB’s «D-» BFSR — which maps to a Baseline [. good franchise as one of Armenia’s largest […] banks (ranking second in terms of assets with a 12.2% market share as at YE2007 — reportedly moving up to first place by March 2008) and good financial metrics, particularly, buoyant profitability, solid capitalisation and above-average efficiency ratios, within the Armenian context. ashib.am |
Мы также добавили черные боковые пороги, 2-дюймовый […] навес, эксклюзивные колеса черного цвета и всесезонные […] более неустрашимого вида. ms-auto.co.jp | We also added black side tube step, 2 inch lift up, exclusive black color […] to make it with a look of fearless determination. ms-auto.co.jp |
Поскольку пропорциональная […] только на термальные нейтроны, полиэтиленовый модератор, […] который замедляет случайные быстрые нейтроны до термальных энергий, окружает нейтронно чувствительную трубу. ru.flukebiomedical.com | Since the BF3 proportional counter […] tube will only respond to thermal neutrons, a polyethylene moderator, which slows the […] incident fast neutrons to thermal energies, surrounds the neutron sensitive tube. flukebiomedical.com |
В настоящий момент компания […] promwad.com | Currently Promwad Company develops a video surveillance and recording system using JPEG2000 image compression standard based on ADV212/202 codec and Blackfin BF561 duo core processor. promwad.com |
SF1605x400 обработанной винт мяч […] zappautomation.co.uk | The SF1605x400 machined ball screw is […] zappautomation.co.uk |
Если заготовка имеет важное значение в стране, то […] составителям кадастров рекомендуется использовать национальные […] ipcc-nggip. | If logging is significant in the […] harvest data or derive country-specific BF values. ipcc-nggip.iges.or.jp |
В январе 2009 года, в рамках ежегодного пересмотра кредитных рейтингов, рейтинговой агентство Moody’s […] подтвердило […] Aa3.ru, что свидетельствует […] о стабильном финансовом положении ОГК-1. ogk1.com | In January 2009 as part of annual revising of credit ratings, the international rating agency Moody’s […] confirmed the international […] an evidence of OGK-1’s stable financial position. ogk1.com |
На устройствах РПН с числом переключений более чем 15.000 в год мы […] рекомендуем применять маслофильтровальную установку OF100 (инструкция по […] сменными фильтрами. highvolt.de | If the number of on-load tap-changer operations per year […] is 15,000 or higher, we recommend the use of […] 100 with a paper filter insert (see Operating Instructions BA 018). highvolt.de |
В нашем […] staubli.com | Discover all the advantages, technical features and part numbers of the SPH/BA couplings in our catalog. staubli.com |
| Запросы и бронирования, связанные с Вознаграждениями (включая Вознаграждения от Компаний-партнеров) можно сделать на сайте ba.com или в местном сервисном центре Участника в соответствии с процедурой оформления Вознаграждений, которая может время от времени быть в силе, как указано на сайте ba.com. britishairways.com | Requests and bookings relating to Rewards (including Service Partner Rewards) may be made online at ba.com or through the Member’s local service centre in accordance with such procedures that may be in force from time to time for the issue of Rewards, as set out on ba.com. britishairways.com |
На грузовики могут устанавливаться зарубежные […] дизели Perkins мощностью 65 л.с. (базовый [. 79 л.с. или отечественный владимирский […] ВМТЗ Д-130Т мощностью 65 л.с. Приводы от валов отбора мощности спереди и сзади позволяют навешивать различное дополнительное оборудование. trucksplanet.com | The trucks can be equipped with foreign […] Perkins 65 hp diesel (Base engine) and Deutz BF 04L 2011 with […] developes 65 hp. trucksplanet.com |
I. Общие сведения о Шанхае должен достичь Фан-Ко, […] ветра предназначены для вентилятора выхлопных […] устройств для удовлетворения оперативных потребностей различных рабочих условиях, он имеет небольшой размер, легкий вес, красивый внешний вид, низкий уровень шума, простота в обслуживании. ru.shyngda.com | I. Overview of Shanghai should reach a Fan Co., the design and […] designed for the blower exhaust devices […] to meet the operational requirements of different working conditions, it has a small size, light weight, beautiful appearance, low noise, easy maintenance. en.shyngda.com |
Наряду со страхованием кредита на инвестиции мы наше предложение расширили на два следующих страховых продукта для страхования […] просроченных задолженностей по экспортным […] банкам откупать экспортные задолженности […] без регресса на экспортера. egap.cz | Simultaneously with insurance of a credit for the financing of investments, we extended our offer by two other insurance products for [. insurance of ceded receivables from export […] banks to purchase export receivables […] without recourse against the exporter. egap.cz |
Оборот […] только на подъемных машинах, превысил миллиард евро (более 15 миллиардов […] эстонских крон) в год. intrac.ee | The turnover of Manitou BF, who is focused […] only on lifting machines, is over one milliard euro (more than 15 milliard Estonian kroons ) a year. intrac.ee |
Для учета коры в изымаемой при заготовке древесине необходимо использовать «долю коры в заготовленной древесине» (BF).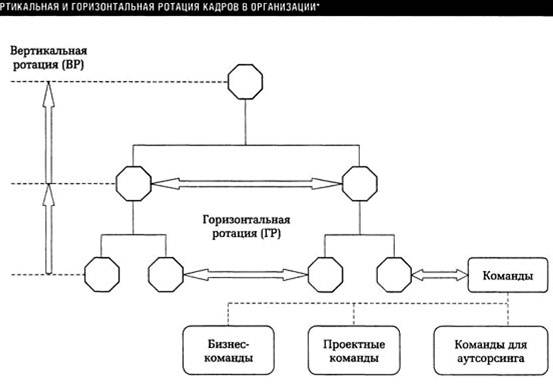 ipcc-nggip.iges.or.jp | Bark fraction in harvested wood (BF) should be 4.33 applied to account for bark in wood removals with harvest. ipcc-nggip.iges.or.jp |
Если бы Володя Малахов, до этого очень здорово […] бы выиграли тот матч, вышли на чистое первое […] место, и, что очень важно, поменялись бы с украинцами местами психологически. crestbook.com | If Volodya Malakhov, who had played that game extremely well until […] then we’d have won the match, moved into […] clear first place and, very importantly, switched places with the Ukrainians psychologically. crestbook.com |
Быстроразъемные […] утечек при разъединении и быстроразъемные полнопоточные соединения DMR для […] систем охлаждения: масляных систем и систем вода/гликоль. staubli.com | SPH/BA clean break and DMR full […] flow quick release couplings for cooling applications such as oil and water glycol connections. staubli.com |
Изъятие древесины (L древ.-изъятия ) рассчитывается с помощью уравнения 2.12 из главы 2, товарные круглые лесоматериалы с корой (H), коэффициент преобразования и […] разрастания биомассы (BCEF ), доля […] к надземной биомассе (R), доля […] углерода в сухом веществе (CF) и табличные данные по умолчанию, раздел 4.5. ipcc-nggip.iges.or.jp | Wood removal (L wood-removals ) is calculated with Equation 2. […] fraction in harvested wood […] fraction of dry matter (CF) […] and default tables, Section 4.5. ipcc-nggip.iges.or.jp |
В Институте агротехники и животноводства Баварского земельного управления сельского хозяйства вот уже много лет […] используются инкубаторы с принудительной […] которым качество исследований остается […] неизменном высоким. binder-world.com | At the Institute for Agricultural Engineering and Animal Husbandry at the Bavarian State Research Center for Agriculture, […] incubators with mechanical convection of the BF [. high quality of research for many years. binder-world.com |
 ..]
..] ..]
..] ..]
..] net
net ..]
..]

 iges.or.jp
iges.or.jp ru, which is […]
ru, which is […] ..]
..]
 ..]
..] ..]
..] 12, Chapter 2, merchantable round wood over bark (H), biomass conversion expansion factor (BCEF ), bark
12, Chapter 2, merchantable round wood over bark (H), biomass conversion expansion factor (BCEF ), bark ..]
..]Кадровые перестановки. Утверждена новая структура Минэнерго
Посты заместителя министра покинули А. Инюцын и А. Тихонов.
Москва, 9 фев — ИА Neftegaz.RU. Министр энергетики РФ Н. Шульгинов утвердил распределение полномочий в министерстве и перечень своих заместителей.
Об этом сообщили в пресс-службе Минэнерго.
На посту замминистра остались:
А. Бондаренко,
Е. Грабчак,
П. Сниккарс,
П. Сорокин,
А. Яновский.
Посты заместителя министра покинули А. Инюцын и А. Тихонов.
Ранее эту должность оставил также Ю. Маневич.
А. Бондаренко будет курировать вопросы законопроектной деятельности (чем она занималась и ранее).
Дополнительно она также будет заниматься вопросами информационного обеспечения, организации делопроизводства и контроля исполнения поручений.
За Е. Грабчаком останется курирование вопросов оперативного управления в ТЭК, техническую политику, вопросы строительства, чрезвычайных ситуаций, топливообеспечения в осенне-зимний период, взаимодействия с регионами.
Е. Грабчак получит новые полномочия в сфере земельных отношений и территориального планирования, развития цифровых технологий (в части ведомства и в сфере электроэнергетики).
В Минэнерго он теперь будет отвечать за государственную информационную систему ТЭК, проектную деятельность и закупки.
П. Сниккарс продолжит курировать сферы электроэнергетики и теплоснабжения.
К его полномочиям добавят регулирование правового обеспечения деятельности министерства и совершенствование регуляторной политики.
П. Сорокин останется курировать нефтегазовый комплекс, стратегическое развитие, госпрограммы, инновации и импортозамещение.
Дополнительно он получит полномочия по курированию цифровой трансформации вопросов экологии и климатической повестки.
А. Яновский все так же будет заведовать вопросами международного сотрудничества, угольной промышленности.
Его новые полномочия коснутся вопросов бюджета министерства, имущественных отношений, координации работы подведомственных организаций и хозяйственного обеспечения.
Напомним, премьер-министр РФ М. Мишустин ранее предложил оптимизировать штат госслужащих.
Сокращение к 1 января 2021 г. должно составить 5% в центральных аппаратах федеральных органов власти и 10% — в территориальных органах.
Освободившиеся средства останутся в фондах оплаты труда.
В мэрии Уфы начались кадровые перестановки :: Башкортостан :: РБК
Фото: сайт мэрии Уфы
В столице Башкирии вице-мэр по строительству Аскат Хайруллин теперь отвечает за транспортную доступность в городе. Статус заместителя главы администрации он сохранил. О новых кадровых решениях объявил временно исполняющий обязанности столичного градоначальника Рустем Газизов на оперативном совещании. В пресс-службе администрации города пояснили РБК Уфа, что перестановки связаны с переходом на другую работу начальника Управления транспорта и связи Ильдара Юланова.
Статус заместителя главы администрации он сохранил. О новых кадровых решениях объявил временно исполняющий обязанности столичного градоначальника Рустем Газизов на оперативном совещании. В пресс-службе администрации города пояснили РБК Уфа, что перестановки связаны с переходом на другую работу начальника Управления транспорта и связи Ильдара Юланова.
Хайруллин пришел работать в мэрию 4 декабря 2018 года. Он стал одним из первых назначенцев Ульфата Мустафина в должности мэра. Родом он из Белебея. Окончил Уфимский государственный нефтяной технический университет и Башкирскую академию государственной службы и управления при Президенте РБ. Трудовую деятельность начинал мастером строительного участка и инженером в тресте, пройдя путь до генерального директора «Башкортостаннефтезаводстроя».
Как сообщал РБК Уфа, 30 ноября депутаты Горсовета Уфы определились с кандидатурой временно исполняющего обязанности главы администрации города. Им стал Рустем Газизов, ранее курировавший вопросы кадров, спорта, молодежи и гражданского общества. Он стал вторым и.о. мэра за месяц по причине временной нетрудоспособности его предшественника Радмила Муслимова, назначенного врио главы администрации 2 ноября.
Он стал вторым и.о. мэра за месяц по причине временной нетрудоспособности его предшественника Радмила Муслимова, назначенного врио главы администрации 2 ноября.
Пост остается вакантным после кончины прежнего мэра Ульфата Мустафина. Чиновник ушел из жизни 29 октября, причиной смерти стали осложнения, возникшие на фоне коронавируса, мэр боролся с болезнью 35 дней, однако медикам не удалось спасти его жизнь.
Автор
Галия Набиева
Перестановки в силовых ведомствах — отставки руководителей 2014
Высокопоставленный свердловский силовик переводится в Магадан. Инсайд
Инсайд
Глава управления охраны общественного порядка ГУ МВД по Свердловской области Олег Тощев готовится к переводу в Магаданскую область. Он претендует на высокий пост в региональном главке полиции, сообщил URA.RU источник в силовых кругах. По версии еще одного инсайдера, в Магадан переедет другой свердловский полицейский — замглавы начальника полиции ГУ МВД по Свердловской области Игорь Алексеев.
21 апреля 2021 12:48
Бастрыкин повысил в должности пермского следователя
Бывшего заместителя руководителя следственного отдела по Индустриальному району Перми Феликса Бочковского повысили в должности. Соответствующий приказ подписал глава Следственного комитета России Александр Бастрыкин, сообщается на сайте СКР по Пермскому краю.
20 апреля 2021 19:23
Свердловская область останется без прокурора еще на месяц
Прокурора Свердловской области в ближайшее время не назначат. Заседание Совета Федерации, на которое может быть вынесен этот вопрос, состоится во второй половине мая. Об этом сообщил источник URA.RU, знакомый с ситуацией.
Заседание Совета Федерации, на которое может быть вынесен этот вопрос, состоится во второй половине мая. Об этом сообщил источник URA.RU, знакомый с ситуацией.
19 апреля 2021 15:29
В прокуратуре Челябинской области новое назначение
Генпрокурор РФ Игорь Краснов подписал приказ о назначении прокурором Уйского района Александра Соловьева. Об этом URA.RU сообщили в пресс-службе прокуратуры Челябинской области. Ранее Соловьев руководил аналогичным ведомством в Чебаркульском районе.
12 апреля 2021 13:18
Бастрыкин устроил разнос свердловскому главку СКР
Председатель Следственного комитета России Александр Бастрыкин провел видеосовещание, в ходе которого подверг критике работу следственного управления СК России по Свердловской области. Информация об этом размещена на официальном сайте СК. По данным высокопоставленных инсайдеров, близких к силовым кругам, времени на исправление ситуации свердловским следователям отвели немного
Информация об этом размещена на официальном сайте СК. По данным высокопоставленных инсайдеров, близких к силовым кругам, времени на исправление ситуации свердловским следователям отвели немного
07 апреля 2021 16:14
Источник: новый прокурор Кургана возьмется за коррупционные дела
Новый прокурор Кургана Виталий Долгушин особое внимание уделит коррупционным делам в городе. Таким мнением с URA.RU поделились инсайдеры из числа бывших сотрудников силовых ведомств.
06 апреля 2021 13:03
В МЧС по Пермскому краю сменилось руководство
Главное управление МЧС по Пермскому краю временно возглавил первый замначальника Игорь Хлебников. Об этом URA.RU сообщили в пресс-службе ведомства.
Об этом URA.RU сообщили в пресс-службе ведомства.
06 апреля 2021 11:25
В Кургане назначили городского прокурора. Инсайд URA.RU подтвердился
Прокурором Кургана стал Виталий Долгушин, экс-прокурор Частоозерского района. Такая информация появилась на сайте областной прокуратуры. Ранее агентство писало, что его кандидатура рассматривалась в Москве.
06 апреля 2021 10:38
В Пермском крае назначен новый замначальника полиции. Инсайд URA.RU подтвердился
Заместителем начальника полиции ГУ МВД России по Пермскому краю назначен Виталий Данилов, сообщили URA.RU в пресс-службе регионального полицейского главка. До этого Данилов возглавлял УМВД России по Чите.
До этого Данилов возглавлял УМВД России по Чите.
02 апреля 2021 16:31
Источник: в пермское ГУ МВД едет третий силовик из Забайкалья
Новым руководителем управления экономической безопасности и противодействия коррупции ГУ МВД по Пермскому краю станет сотрудник забайкальского УМВД. Источник URA.RU в силовых структурах рассказал, что этот сотрудник станет уже третьим руководителем в полицейском главке Прикамья, который за последние месяцы перевелся в регион с Дальнего Востока.
31 марта 2021 08:46
Инсайд: назначен замначальника полиции Пермского края
Заместителем начальника полиции ГУ МВД России по Пермскому краю назначен силовик из Забайкальского края, рассказал URA. RU источник в полицейском главке региона. Предполагается, что экс-начальник УМВД по Чите Виталий Данилов занял в Перми должность, которую ранее занимал начальник УМВД по Забайкалью Вячеслав Еговцев.
RU источник в полицейском главке региона. Предполагается, что экс-начальник УМВД по Чите Виталий Данилов занял в Перми должность, которую ранее занимал начальник УМВД по Забайкалью Вячеслав Еговцев.
29 марта 2021 20:51
Источник: шеф пермского Центра «Э» уходит из МВД
Руководитель центра по противодействию экстремизму ГУ МВД России по Пермскому краю полковник полиции Евгений Иванов уйдет в отставку. Об этом URA.RU рассказал источник, знакомый с ситуацией. Место Иванова может занять начальник краевого управления экономической безопасности и противодействия коррупции полковник полиции Александр Журахов.
26 марта 2021 20:48
Путин назначил нового прокурора ЯНАО
Виктор Эпп назначен на должность прокурора ЯНАО. Указ подписал президент РФ Владимир Путин 24 марта. Документ опубликован на официальном интернет-портале правовой информации.
Указ подписал президент РФ Владимир Путин 24 марта. Документ опубликован на официальном интернет-портале правовой информации.
24 марта 2021 20:14
Силовик из столицы ЯНАО стал шефом полиции Нового Уренгоя
Подполковника полиции Дмитрия Соловьева назначили руководителем ОМВД по Новому Уренгою (ЯНАО). До вчерашнего дня офицер возглавлял отдел МВД России по Салехарду, сообщается на официальном сайте ведомства.
23 марта 2021 15:10
Источник: в свердловской прокуратуре ждут отставку руководителя
Врио прокурора Свердловской области Владимир Маленьких ушел в отпуск, из которого может не вернуться. По данным источника, близкого к уральской прокуратуре, Маленьких планирует написать рапорт об отставке.
23 марта 2021 12:56
Путин провел масштабные перестановки среди силовиков
Президент России Владимир Путин снял с должностей несколько высокопоставленных сотрудников силовых ведомств. Соответствующий документ имеется в распоряжении URA.RU.
19 марта 2021 13:10
Путин отправил в отставку главу МЧС по Пермскому краю
Начальник главного управления МЧС России по Пермскому краю Андрей Бабинцев освобожден от занимаемой должности. Соответствующий указ №149 от 18 марта подписал президент РФ Владимир Путин (есть в распоряжении URA.RU).
19 марта 2021 10:18
Новый начальник челябинского УФСБ получил важный пост
Губернатор Челябинской области Алексей Текслер включил в состав координационного совещания по обеспечению правопорядка в регионе нового начальника УФСБ Дмитрия Иванова. Одновременно места в нем потеряли предшественник Иванова Сергей Сизов и экс-прокурор региона Виталий Лопин. Соответствующее распоряжение опубликовано на интернет-портале официальной информации.
Одновременно места в нем потеряли предшественник Иванова Сергей Сизов и экс-прокурор региона Виталий Лопин. Соответствующее распоряжение опубликовано на интернет-портале официальной информации.
18 марта 2021 13:37
Источник: новый прокурор приедет в ЯНАО за погонами и пенсией
Для будущего прокурора ЯНАО Виктора Эппа назначение станет возможностью получить «генеральские» погоны и уйти на пенсию с соответствующими выплатами. Такую версию развития событий обсуждают сотрудники надзорных органов, сообщают источники URA.RU в силовых структурах.
17 марта 2021 07:42
Будущий челябинский прокурор отдавал под суд карельских VIP. Чего ждать от нового руководителя
Будущий прокурор Челябинской области Карен Габриелян на посту прокурора Республики Карелия подписывал обвинительные заключения в отношении руководителей самого высокого уровня. Журналисты из Карелии рассказали URA.RU, чем запомнился Габриелян местной прессе.
Журналисты из Карелии рассказали URA.RU, чем запомнился Габриелян местной прессе.
15 марта 2021 15:47
Совфеду представилась возможность оценить перестановку региональных прокуроров
Фото: council.gov.ru
В Совет Федерации поступило представление президента России по кандидатурам на должности прокуроров субъектов страны, сообщили в пресс-службе СФ. Новая процедура проходит во второй раз в соответствии с поправками к Конституции РФ.
«Представление президента РФ было направлено в Совет Федерации в соответствии с частью 4 статьи 129 Конституции России и пунктом 11 статьи 151 Федерального закона «О прокуратуре Российской Федерации», — отметил председатель комитета СФ по конституционному законодательству и государственному строительству Андрей Клишас.
По его словам, в соответствии с регламентом комитеты Совфеда проведут необходимые консультации и направят консолидированную позицию по представленным кандидатурам главе СФ.
Консультации в верхней палате парламента пройдут 9 февраля. Уже определены претенденты. Так, в ряде субъектов РФ намечена ротация руководящих кадров.
Прокурора Кемеровской области Павла Бухтоярова предлагается перевести на аналогичную должность в Пермский край. Прокурора Тульской области Романа Праскова — в Ростовскую область. Прокурора Ивановской области Андрея Ханько — в Иркутскую область. Прокурора Мурманской области Василия Щербакова — в Республику Мордовия.
«В соответствии с Конституцией России Президент Российской Федерации назначает на должность после консультаций с Советом Федерации и освобождает от должности, в частности, прокуроров субъектов РФ, прокуроров военных и других специализированных прокуратур, приравненных к прокурорам субъектов РФ», — напомнили в пресс-службе СФ.
Как рассказывал L.R, после внесения изменений в законодательство прокуроры субъектов Федерации и приравненные к ним прокуроры будут назначаться на должность президентом после консультаций с Советом Федерации, а освобождаться от должности исключительно президентом. Ранее назначение и освобождение генпрокурора на должность осуществлял Совфед по представлению президента. А прокуроры субъектов назначались президентом по представлению генпрокурора, согласованному с субъектом РФ.
Ранее назначение и освобождение генпрокурора на должность осуществлял Совфед по представлению президента. А прокуроры субъектов назначались президентом по представлению генпрокурора, согласованному с субъектом РФ.
Для консультации по кандидатуре на должность генпрокурора президент направляет в Совет Федерации представление и другие материалы, характеризующие кандидата. При этом Совфед не позднее недельного срока со дня получения материалов информирует в письменной форме главу государства о результатах рассмотрения.
В декабре 2020 года в Совфеде прошла первая подобная консультация после принятия поправок из «конституционного пакета». Тогда прокурором Амурской области президент предложил назначить Александра Бучмана, прокурором Тамбовской области — Александра Гулягина, прокурором Оренбургской области — Руслана Медведева, прокурором Воронежской области — Николая Савруна, прокурором Республики Хакасия — Сергея Фирсова. На должность Дальневосточного транспортного прокурора предлагается назначить Александра Воронина, на должность Западно-Сибирского транспортного прокурора — Илью Сухоносова.
Первые перестановки Дегтярева в Хабаровске: оценка
Временный глава Хабаровского края Михаил Дегтярев сделал первые кадровые замены. Свои места покинули два зампреда правительства и советник губернатора. По просьбе «360» политологи рассказали, насколько верными стали перестановки в такое неспокойное для региона время.
Михаил Дегтярев провел первые кадровые перестановки в руководстве Хабаровского края. Свой пост покинул первый заместитель председателя правительства, руководитель аппарата Владимир Хлапов. Его место займет коллега Дегтярева по Госдуме Александр Никитин. Ушел и советник губернатора Захар Синяговский. В ближайшее время, уточнил временный губернатор во время очередного Instagram-эфира, он подпишет заявление на увольнение первого заместителя председателя правительства по экономическим вопросам Юрия Золочевского.
Экс-губернатора края Сергея Фургала отправили в отставку после ареста по подозрению в организации убийств и покушении на бизнесменов. По словам председателя СК Александра Бастрыкина, следователи собрали серьезные доказательства причастности Фургала к преступлениям.
Осторожный минимум
Пока Дегтярев сделал минимальные перестановки, пояснил «360» вице-президент Центра политических технологий Алексей Макаркин. Заменил руководителя аппарата — самого близкого к губернатору человека. Сейчас время для больших замен не вполне удачное из-за холодного приема в Хабаровске.
«Что касается Золочевского, непонятно, то ли Дегтярев его уволил, то ли он сам решил уйти. Это пока непонятно, равно как непонятно, кем его Дегтярев заменит. Если свой руководитель аппарата у него есть, есть ли кандидат на должность первого зама — пока большой вопрос», — добавил Макаркин.
22 июля временный губернатор сообщил, что планирует работать с командой своего предшественника. Но в любом случае, уверен политолог, в правительстве края будут и дальше происходить изменения, но это будет зависеть от того, сумеет ли новый глава выстроить отношения с местными жителями и сможет ли усилить свои позиции в регионе.
«Если Дегтярев сейчас утвердится, то могут заменить достаточно многих. Потому что глава региона, который приходит, обычно проводит свою кадровую политику. Но это будет зависеть от того, какие у него будут позиции в регионе. Люди, которые будут приходить [в его команду], будут исходить из того, какие позиции у действующего губернатора, какой авторитет среди населения», — заключил политолог.
Потому что глава региона, который приходит, обычно проводит свою кадровую политику. Но это будет зависеть от того, какие у него будут позиции в регионе. Люди, которые будут приходить [в его команду], будут исходить из того, какие позиции у действующего губернатора, какой авторитет среди населения», — заключил политолог.
Со своим уставом
Политтехнолог Наталия Елисеева напомнила, что Дегтярев не планировал менять старую команду. Он был назначен временно исполняющим обязанности, то есть его присутствие не предполагало кадровых перестановок. Даже если причины для смены были, с этим стоило бы немного подождать. Время выбрано неудачно.
«В данном случае перестановка кадров может быть расценена как наведение своих порядков. Что может усугубить и так противоречивую ситуацию в регионе. То есть, опять же, вместо того чтобы как-то налаживать ситуацию, утихомирить, мы наблюдаем посредством кадровых перестановок очередной повод для недовольства людей», — пояснила Елисеева «360».
Для начала стоило выстроить позитивный имидж, наладить живой канал взаимодействия с жителями. А если люди видят губернатора в интернете, в салоне машины бизнес-класса с наушником в ухе, коммуникации на пользу не идет.
А если люди видят губернатора в интернете, в салоне машины бизнес-класса с наушником в ухе, коммуникации на пользу не идет.
«А в регионе есть запрос на коммуникацию, на простоту. В сравнении с бывшим губернатором Сергеем Фургалом Дегтярев в плане позиционирования проигрывает. Потому что Фургал в этом плане был попроще», — отметила она.
В целом смена двух заместителей и советника довольно предсказуема. Регион проблемный, есть сложности с инвестициями, развитием края. В принципе, кадровое обновление требовалось. Уволив тех, кто курировал эти вопросы, Дегтярев начал пытаться исправить ситуацию.
«Скорее всего, будет сменена большая часть команды. Новый управленец подбирает людей под себя, под свой стиль управления и те вопросы, которые стоят перед ним и регионом. С учетом [установки] рубить с плеча с самого начала, думаю, на следующей неделе мы увидим ряд замен», — сказала Елисеева.
Бета-транскриптов Т-клеточного рецептора вне кадра элиминируются множественными путями in vivo
Abstract
Непродуктивные гены антигенных рецепторов со сдвигами рамки считывания, возникающими во время сборки этих генов, обнаруживаются во многих зрелых лимфоцитах. Транскрипты этих генов имеют кодоны преждевременной терминации (PTC) и могут кодировать усеченные белки, если они не инактивированы или не уничтожены нонсенс-опосредованным распадом (NMD). В клетках млекопитающих NMD может активироваться путями, которые зависят от присутствия интрона, расположенного ниже PTC; однако NMD также может быть активирован путями, которые не зависят от этих нижестоящих интронов, а пути, независимые от NMD, могут инактивировать PTC-содержащие транскрипты.Здесь, путем создания и анализа мышей с генно-направленными модификациями локуса эндогенного Т-клеточного рецептора бета ( Tcrb ), мы демонстрируем, что в Т-клетках in vivo оптимальный клиренс PTC-содержащих транскриптов Tcrb зависит от на наличие интрона ниже PTC.
Образец цитирования: Mahowald GK, Mahowald MA, Moon C, Khor B, Sleckman BP (2011) Бета-транскрипты рецепторов Т-клеток вне кадра устраняются множественными путями In Vivo .PLoS ONE 6 (7):
e21627.
https://doi.org/10.1371/journal.pone.0021627
Редактор: Себастьян Д. Фугманн, Национальный институт старения, Соединенные Штаты Америки
Поступила: 29 апреля 2011 г .; Одобрена: 3 июня 2011 г .; Опубликовано: 13 июля 2011 г.
Авторские права: © 2011 Mahowald et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа поддержана грантом AI47829 Национальных институтов здравоохранения (NIH). Мыши были произведены в центре трансгенных заболеваний, поддерживаемом Центральным центром ревматических заболеваний при Вашингтонском университете (NIH P30-AR48335), и размещены в учреждении, поддерживаемом Национальным центром исследовательских ресурсов (NCRR), грант RR012466. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Цепи рецептора антигена лимфоцитов состоят из N-концевых вариабельных областей, кодируемых первыми двумя экзонами гена рецептора антигена, и константных областей C-констант, кодируемых остальными экзонами [1]. Второй экзон всех генов рецепторов лимфоцитарного антигена собирается посредством реакции рекомбинации V (D) J, которая инициируется, когда эндонуклеаза RAG вводит двухцепочечные разрывы ДНК на границе переменной рекомбинации (V), разнообразия (D) и соединения ( J) генные сегменты и их фланкирующие последовательности узнавания RAG (сигналы рекомбинации, RS) [2].Эти двухцепочечные разрывы ДНК репарируются с помощью пути негомологичного соединения концов (NHEJ), соединяющего генные сегменты V, D и J [1]. NHEJ неточен, и случайное накопление и потеря нуклеотидов, которое сопровождает этот процесс соединения, имеет важное значение для диверсификации гена рецептора антигена и адаптивного иммунитета. Однако такая диверсификация также приводит к сдвигам рамки считывания и кодонам преждевременной терминации (PTC) в двух из трех рамок считывания. Эти аллели рецептора антигена вне рамки активно транскрибируются в лимфоцитах, обеспечивая богатый источник PTC-содержащих транскриптов, которые, если не будут инактивированы или уничтожены нонсенс-опосредованным распадом (NMD), могут привести к продукции усеченных пептидов рецептора антигена, которые могут быть вредным для развивающихся лимфоцитов.В соответствии с этим представлением избирательное удаление Upf2, центрального медиатора NMD, в тимоцитах приводит к дефектам в развитии Т-клеток, что может быть частично связано с накоплением транскриптов генов рецепторов Т-клеток с кодонами преждевременной терминации [3] .
У низших эукариот NMD активируется, когда PTC распознаются на основании их расстояния от поли (A) тракта [4]. Напротив, клетки млекопитающих могут активировать NMD через пути, которые распознают терминирующие кодоны как преждевременные, если они лежат выше вырезанного интрона, независимо от их расстояния от поли (A) тракта [5], [6], [7], [8] ].Активация этого пути, называемого EJC-зависимым NMD, требует, чтобы комплексы соединений экзонов (EJCs) существовали ниже рибосом, остановленных на PTCs во время первого раунда трансляции [8], [9]. EJC откладываются на 20–24 н. Выше границы экзон-экзон, и для того, чтобы PTC активировал NMD посредством EJC-зависимых путей, он должен располагаться на расстоянии более 50-55 п.н. перед экзон-экзоном. соединение [7], [9], [10], [11], [12].
Транскрипты, которые имеют PTC, расположенные менее чем на 50 п.н. выше соединения экзон-экзон, или которые имеют PTC в последнем экзоне, все еще способны активировать NMD в клетках млекопитающих [5], [11], [12], [13], [14].Полагают, что активация NMD этим безотказным путем требует определения расстояния PTC от поли-A тракта, как в случае с низшими эукариотами [15]. Хотя активация отказоустойчивой NMD не зависит от присутствия интрона ниже PTC, она требует присутствия интрона в другом месте гена [11], [14], [16], [17], [18] ]. Кроме того, механизмы, которые, например, изменяют сплайсинг, могут инактивировать PTC-содержащие транскрипты, не приводя к их деградации [19], [20], [21], [22], [23], [24], [25], [26].В связи с этим пре-мРНК с PTC могут накапливаться в ядре без сплайсинга или сплайсинга не полностью. Более того, транскрипты, содержащие PTC, можно альтернативно сплайсировать, удаляя экзон с PTC, этот процесс называется нонсенс-ассоциированным измененным сплайсингом. Как PTC воспринимаются в этих не полностью обработанных транскриптах, полностью не изучено. Вместе эти механизмы предотвращают кодирование транскриптами с PTC усеченных белков, которые могут иметь пагубные последствия.
Транскрипты локуса цепи бета-рецептора Т-клеток
( Tcrb ), содержащие PTC, легко разрушаются NMD in vivo [27].Здесь мы разрабатываем подход для непосредственного определения потребности в интроне, расположенном ниже PTC для эффективного очищения транскриптов, шаблонизированных эндогенным локусом Tcrb в тимоцитах. Используя многоступенчатое нацеливание на гены, мы создали две минимально модифицированные версии локуса Tcrb мыши, Tcrb A и Tcrb F , которые претерпевают нормальную реаранжировку и экспрессию в развивающихся тимоцитах. Эти два аллеля идентичны, за исключением того, что PTC аллеля Tcrb A будут иметь нижележащие интроны, тогда как PTC аллеля Tcrb F не будут.Сравнивая стабильность PTC-содержащих транскриптов Tcrb A и Tcrb F в тимоцитах, мы демонстрируем, что нормальный клиренс этих транскриптов зависит от присутствия интронов ниже PTC.
Результаты и обсуждение
Генерация
Tcrb A и Tcrb F аллелей
Локус мыши Tcrb занимает 0,7 Мб, с 34 сегментами гена Vb, лежащими в 0.Область 4 Mb перед двумя кластерами Db-Jb, каждый с одним Db и 6 или 7 сегментами гена Jb (Fig. 1a) [28]. Второй экзон завершается, когда сегмент гена Vb перестраивается в реаранжировку DJb в любом из кластеров Db-Jb. Четыре экзона константной области лежат ниже каждого кластера Db-Jb (Cb1 и Cb2) (рис. 1a). Перестройки VDJb в DJb1 транскрибируются с четырьмя экзонами Cb1; аналогично реаранжировки VDJb в DJb2 транскрибируются с четырьмя экзонами Cb2.
Рисунок 1. Аллели Tcrb A и Tcrb F .
( a ) Схема локуса мыши Tcrb . Показаны сегменты генов Vb, Db и Jb (открытые прямоугольники), RS (открытые треугольники), экзоны Cb1 и Cb2 (синие прямоугольники), энхансеры (Eb и промоторы (белые кружки) и последовательностей loxP (заштрихованные овалы). () b ) Схема аллелей Tcrb A и Tcrb F .
https://doi.org/10.1371/journal.pone.0021627.g001
Многоступенчатое нацеливание на гены было использовано для создания мышей с двумя модифицированными эндогенными локусами Tcrb ( Tcrb A и Tcrb F , рис.1б). Первоначально сегмент гена Db1 был удален, ограничивая перестройку сегментов гена Vb кластером сегментов гена Db2-Jb2 (этап A, рис. 1a, см. Аллель Jb1 M3 в ссылке [28]). Сегменты гена Db2, Jb2.1, Jb2.2 и Jb2.3 затем были заменены реаранжировкой DJb2.3 в его нативной конфигурации с образованием аллеля Tcrb A (этап B, фиг. 1a и S1). Аллель Tcrb F идентичен аллелю Tcrb A , за исключением того, что четыре экзона Cb2 были заменены фрагментом ДНК, содержащим слияние этих экзонов без каких-либо интронов (этап C, рис.1а и S2). Важно отметить, что аллели Tcrb A и Tcrb F были созданы посредством умеренных генно-направленных модификаций эндогенного локуса Tcrb дикого типа, аллелей Tcrb A и Tcrb F остаются под той же регуляцией cis (эндогенные промоторы и энхансеры Tcrb ), что и локус Tcrb дикого типа.
Развитие Т-клеток у
Tcrb A / A и Tcrb F / F мышей
Эффективная сборка и экспрессия продуктивного гена цепи Tcrb необходимы для нормального развития Т-клеток.Несколько линий доказательств демонстрируют, что реаранжировка аллелей Tcrb A и Tcrb F происходит с почти нормальной эффективностью. Во-первых, в лимфатических узлах Tcrb A / + и Tcrb F / + Т-клеточных гибридом, полная перестройка происходила с высокой частотой как на аллелях Tcrb A , так и на Tcrb F . (Рис. 2а и Таблица S1). Более того, анализ Tcrb A / A и Tcrb F / F Т-клеточных гибридом показал, что 32% и 35% соответственно имеют полные перестройки VDJb на обоих аллелях (рис.2b и таблица S2). Это близко к максимальным 40%, ожидаемым из-за исключения аллелей [29], [30]. Наконец, анализ Tcrb A / F Т-клеточных гибридом с одиночными реаранжировками VDJb выявил примерно равные частоты реаранжировок VDJb на аллелях Tcrb A и Tcrb F (рис. 2c и таблица S3). .
Рисунок 2. Развитие Т-клеток у мышей Tcrb A / A и Tcrb F / F .
( a ) Процент аллелей Tcrb A (красный), Tcrb F (зеленый) и Tcrb + (синий) в конфигурации VDJb в Tcrb A / + и Tcrb F / + гибридомы. ( b ) Процент гибридом Tcrb A / A и Tcrb F / F с одним (синий) или двумя (красный) полностью реаранжированными аллелями. ( c ) Процент гибридом Tcrb A / F с единственной полной реаранжировкой VDJb на аллеле Tcrb A (синий) или Tcrb F (красный).( d ) Проточно-цитометрический анализ экспрессии CD4 и CD8 в тимоцитах мышей Tcrb + / + , Tcrb A / A и Tcrb F / F . Указаны проценты. Данные являются репрезентативными по крайней мере для 4 мышей, проанализированных для каждого генотипа. ( e – f ) Количество тимоцитов DN, DP, CD4SP и CD8SP (e) и количество CD4 + и CD8 + Т-лимфоцитов селезенки (f) в Tcrb + / + (синий) , Tcrb A / A (красный) и Tcrb F / F (зеленый) мышей, изображенных как среднее +/- SEM.По каждому генотипу анализировали не менее четырех мышей соответствующего возраста.
https://doi.org/10.1371/journal.pone.0021627.g002
Развитие тимоцитов у мышей Tcrb A / A и Tcrb F / F было неотличимо от мышей дикого типа (рис. 2d). и е). В связи с этим, мыши дикого типа ( Tcrb + / + ), Tcrb A / A и Tcrb F / F имели схожее количество CD4 — : CD8 — (двойной отрицательный, DN), CD4 + : CD8 + (двойной положительный, DP) и CD4 — : CD8 + или CD4 + : CD8 — (одиночный положительный, SP) тимоцитов (рис.2г и д). Проточно-цитометрический анализ экспрессии цепи Tcrb не выявил значимых различий между тимоцитами Tcrb A / A , Tcrb F / F и Tcrb + / + (данные не показаны). Наконец, мыши Tcrb A / A , Tcrb F / F и Tcrb + / + имеют одинаковое количество зрелых CD4 + и CD8 + Т-клеток селезенки (рис. 2f ). Взятые вместе, эти данные демонстрируют, что аллели Tcrb A и Tcrb F эффективно реаранжируются и экспрессируются и могут поддерживать нормальное развитие Т-клеток.
Дифференциальная стабильность PTC-содержащих
Tcrb A и Tcrb F транскриптов
Подобно локусу Tcrb дикого типа, Tcrb A PTC лежат в третьем из шести экзонов, тогда как Tcrb F PTC лежат в третьем и последнем экзоне; таким образом, Tcrb A PTCs имеют нижележащие интроны, тогда как Tcrb F PTC не имеют (рис. 3a). Примечательно, что PTC расшифровки Tcrb A и Tcrb F находятся на одинаковом расстоянии (0.7 т.п.н.) из поли (А) тракта и имеют два вышележащих интрона в тех же местах (рис. 3а). Таким образом, сравнение стабильности PTC-содержащих Tcrb A и Tcrb F транскриптов в Tcrb A / A и Tcrb F / F тимоцитов позволяет определить относительный вклад механизмы, которые полагаются на интроны ниже PTC в опосредовании деградации PTC-содержащих транскриптов Tcrb in vivo .
Рисунок 3. PTC-содержащие Tcrb F транскрипты проявляют устойчивость к NMD.
( a ) Tcrb A и Tcrb F аллелей с полностью собранными вторыми экзонами (VDJb), с четырьмя экзонами Cb2 (синие прямоугольники 3, 4 5 и 6, Tcrb A ) или слияние этих экзонов (синий прямоугольник 3, Tcrb F ). Показаны положения двух потенциальных PTC (красные восьмиугольники) и нормального терминирующего кодона (TC, открытый восьмиугольник).Показаны пре-мРНК с экзонами (зеленые столбцы) и интроны (соединяющие черные линии), а также полностью процессированные PTC-содержащие Tcrb A и Tcrb F транскрипты (зеленые столбцы) с соединениями экзонов (EJ, фиолетовые точки). ( b ) Снижение кратности (относительно PTC-содержащих аллелей) PTC-содержащей пре-мРНК Tcrb (светлые столбцы) и полностью процессированной мРНК (красные столбцы) в Tcrb A / A и Tcrb Ф / Ф ДН тимоцитов.P-значения были рассчитаны с помощью биномиального теста для пре-мРНК по сравнению с мРНК и с помощью моделирования Монте-Карло для Tcrb A / A по сравнению с Tcrb F / F .
https://doi.org/10.1371/journal.pone.0021627.g003
С этой целью тимоциты Tcrb A / A и Tcrb F / F CD25 + DN были очищены потоком. цитометрическая сортировка клеток и перестройки VDJb с использованием пяти различных сегментов гена Vb были амплифицированы и секвенированы из геномной ДНК и кДНК, полученных как из пре-мРНК Tcrb , так и из зрелых транскриптов.Всего было проанализировано 1592 последовательности для идентификации тех, которые находятся в кадре (без PTC, PTC —) и тех, которые находятся вне кадра (PTC-содержащие, PTC + ) (Таблица S4). По сравнению с геномной ДНК, фракция пре-мРНК PTC + Tcrb была аналогична доле аллелей PTC + Tcrb как в Tcrb A / A , так и в Tcrb F / F. DN тимоцитов (рис. 3b, светлые столбцы и таблица S4). По сравнению с геномной ДНК наблюдается уменьшение в 12 раз количества мРНК PTC + зрелых Tcrb A (рис.3b, красная полоса и таблица S4). В противоположность этому, количество мРНК PTC + Tcrb F снижается только в три раза (рис. 3b, красная полоса и таблица S4). Таким образом, содержащие PTC мРНК Tcrb A элиминируются более эффективно, чем содержащие PTC мРНК Tcrb F .
Заключительные замечания
Здесь мы показываем, что PTC-содержащие Tcrb A и Tcrb F транскрипты обладают разными способностями к элиминации в тимоцитах млекопитающих in vivo .Единственное различие между аллелями Tcrb A и Tcrb F заключается в наличии интронов, расположенных ниже PTC. Таким образом, эти находки демонстрируют, что эти нижестоящие интроны являются механически важными компонентами в эффективном устранении PTC-содержащих Tcrb транскриптов in vivo , что согласуется с представлением о том, что они необходимы для активации EJC-зависимой NMD. Тем не менее, количество PTC-содержащих транскриптов, шаблонизированных аллелем Tcrb F , сокращается в три раза.Таким образом, механизмы, которые не зависят от нижестоящих интронов, также способны устранять PTC-содержащие транскрипты, хотя и не до уровней, достигаемых при наличии нижестоящих интронов. Поскольку транскрипты, созданные по шаблону аллеля Tcrb F , будут иметь два интрона, возможно, PTC-содержащие Tcrb F транскрипты также могут быть инактивированы нонсенс-ассоциированным измененным сплайсингом или другими механизмами, которые изменяют сплайсинг PCT-содержащих стенограммы. Примечательно, что PTC в гене каппа легкой цепи эндогенного иммуноглобулина часто находятся в последнем экзоне, и, как транскрипты Tcrb F в тимоцитах, количество содержащих PTC каппа-транскриптов легкой цепи иммуноглобулина также снижается примерно в три раза в развивающиеся В-клетки [19], [31].Мы пришли к выводу, что хотя PTC-содержащие транскрипты Tcrb могут элиминироваться с помощью нескольких механизмов в развитии лимфоцитов in vivo , максимальная элиминация этих транскриптов зависит от механизмов, которые зависят от присутствия интронов ниже PTC.
Материалы и методы
Заявление об этике
Это исследование было проведено в строгом соответствии с рекомендациями Руководства по уходу и использованию лабораторных животных Национального института здоровья.Протокол был одобрен Комитетом по изучению животных Вашингтонского университета (№ 20070189).
Поколение
Tcrb A / A и Tcrb F / F мышей
Плечо 5′-гомологии pLNTK-DJ (рис. S1) содержит реаранжировку Db2Jb2.3, амплифицированную с помощью 5’HDJ и 3’Jb2-3, и сегменты гена Jb2.4-Jb2.7, амплифицированные с помощью 5’Jb2.4US. и 3’Jb2.7DS (олигонуклеотидные последовательности перечислены в таблице S5). Сегмент гена Jb2.3 имеет единственную замену G на C для устранения PTC, созданного с использованием 5’Jb2.3, Jb2.3M1, Jb2.3M2 и 3’Jb2.3X, олигонуклеотиды. Плечо 3′-гомологии представляет собой фрагмент ClaI / SpeI в 3,2 т.п.н. ниже Jb2.7. Плечо 5′-гомологии pLNTK-Cb2F (фиг. S2) представляет собой фрагмент ClaI / PstI Tcrb размером 2 т.п.н. Плечо 3′-гомологии было создано путем амплификации константной области кДНК Tcrb с олигонуклеотидами от A до F, 5’Cb2-3’H и 3’Cb2-3’H, как показано на рисунке S2. Делеция сегмента гена Db1 описана в [32]. Эмбриональные стволовые клетки (ЭС) трансфицировали, отбирали и вводили в бластоцисты C57BL / 6, как описано ранее [32].Скрещивание мышей Tcrb A / + и Tcrb F / + привело к ожидаемым менделевским отношениям Tcrb A / A и Tcrb F / F мышей соответственно.
Саузерн-блоттинг
Саузерн-блот-анализ ES-клеток, нацеленных на pLNTK-DJ, проводили на SacI -расщепленной геномной ДНК с зондом А, как описано ранее [33]. Для ES-клеток, нацеленных с помощью pLNTK-Cb2F, расщепленную геномную ДНК PstI зондировали с помощью геномного фрагмента HincII / BamHI длиной 400 п.о. .
Гибридомы
Было создано
гибридом и проанализировано реаранжировок гена Tcrb , как описано ранее [23].
Проточно-цитометрические анализы и очистка клеток
Проточные цитометрические анализы были выполнены на FACSCalibur (BD Biosciences) с использованием FITC-конъюгированного анти-CD25, PE-Cy5-конъюгированного анти-CD4 и FITC-конъюгированного анти-CD8. CD25 + DN тимоциты очищали от 4–5 мышей для каждого генотипа методом проточной цитометрической сортировки клеток (FACSVantage BD Biosciences).
Анализ последовательности
Геномную ДНК и РНК выделяли, как описано ранее, из тимоцитов CD25 + DN, очищенных с помощью проточной цитометрической сортировки клеток [33], [34]. Набор для обратной транскриптазы SuperScriptII (Invitrogen) использовали для синтеза кДНК с использованием олиго-dT или интронного олигонуклеотида 3’Jb2-11 для зрелой мРНК или пре-мРНК, соответственно. Перестройки VDJb амплифицировали из ДНК с использованием Vb2, Vb6, Vb8.1, Vb14 или Vb16 и 3’Jb2-6. Условиями ПЦР были 200 нг геномной ДНК в 50 мкл с 1 мМ MgCl 2 , 100 мМ dNTP и 10 пикомолей каждого олигонуклеотида в течение 17 циклов при 92 ° C (1–00), 60 ° C (1–30) и 72 ° C (1–30) с последующей вторичной амплификацией в течение 30 циклов с использованием 3’Jb2-3 и того же олигонуклеотида Vb.кДНК амплифицировали, как указано выше, за исключением использования олигонуклеотидов Cb2-1 и Cb2-2 в первичной и вторичной реакциях соответственно. Достоверность кратного уменьшения пре-мРНК или мРНК по сравнению с ДНК рассчитывалась с использованием биномиального теста. Значение p для кратного уменьшения мРНК для Tcrb A по сравнению с Tcrb F было рассчитано с использованием моделирования Монте-Карло (сценарий доступен по запросу).
Дополнительная информация
Рисунок S1.
Стратегия таргетинга для генерации аллеля Tcrb A . Генерация аллеля Tcrb A . Показана схема части аллеля Tcrb , в которой сегмент гена Db1 был удален (вверху, Jb1 M3 в ссылке [28]). Сегменты генов Db и Jb показаны как открытые прямоугольники (за исключением Jb2.3, показанного как заштрихованный прямоугольник), а RS — как открытые треугольники. Также показаны экзоны Cb1 и Cb2 (синие прямоугольники), как и целевой вектор pLNTK-DJ, используемый для генерации аллеля Tcrb A + Neo , который имеет целевую замену Db2, Jb2.1, сегменты гена Jb2.2 и Jb2.3 с реаранжировкой Db2Jb2.3 и фланкированный loxP ген устойчивости к неомицину (Neo R ). Показан аллель Tcrb A , образованный после Cre-опосредованной делеции гена устойчивости к неомицину, оставляющий единственный сайт loxP (закрашенный овал). Показаны относительные положения различных сайтов рестрикции, как и зонд А, который использовали для саузерн-блот-анализа различных целевых аллелей. Также показан Саузерн-блот геномной ДНК из целевых линий ES-клеток, переваренных SacI и гибридизированных с зондом A.Указаны маркеры молекулярной массы и относительные положения полос, генерируемых различными аллелями Tcrb .
https://doi.org/10.1371/journal.pone.0021627.s001
(TIF)
Рисунок S2.
Стратегия таргетинга для генерации аллеля Tcrb F . a ) Генерация сплава Cb2. Показаны схемы четырех экзонов Cb2 (помечены с 1 по 4) в геномной ДНК и в кДНК, полученной из полностью процессированной мРНК, и олигонуклеотидов от A до F (обозначены стрелками). b ) Генерация аллеля Tcrb F . Показана схема области Cb2 аллеля Tcrb A (вверху) и вектора таргетинга pLNTK-Cb2F, используемых для создания аллеля Tcrb F + Neo , который имеет целевую замену четырех Cb2 экзоны с фрагментом ДНК, содержащим слияние этих экзонов и гена устойчивости к неомицину, фланкированного loxP-. Также показан аллель Tcrb F , образованный после Cre-опосредованной делеции гена устойчивости к неомицину, оставляя единственный сайт loxP .Показаны относительные положения различных сайтов рестрикции, а также зонды A и B, которые использовали для саузерн-блоттинга. Показаны саузерн-блоты геномной ДНК из целевых линий ES-клеток, которые были расщеплены PstI и гибридизированы с зондом B или расщеплены SacI и гибридизированы с зондом A. Указаны маркеры молекулярной массы и относительное положение полос, генерируемых различными аллелями Tcrb .
https://doi.org/10.1371 / journal.pone.0021627.s002
(TIF)
Благодарности
Мы благодарим доктора Юджина Ольца за критический обзор рукописи.
Вклад авторов
Задумал и спроектировал эксперименты: BPS GKM BK. Проведены эксперименты: ГКМ МАМ СМ. Проанализированы данные: ГКМ МАМ БПС. Написал бумагу: БПС ГКМ.
Ссылки
- 1.
Bassing CH, Swat W, Alt FW (2002) Механизм и регуляция хромосомной рекомбинации V (D) J.Ячейка 109: SupplS45–55. - 2.
Fugmann SD, Lee AI, Shockett PE, Villey IJ, Schatz DG (2000) Белки RAG и рекомбинация V (D) J: комплексы, концы и транспозиция. Анну Рев Иммунол 18: 495–527. - 3.
Weischenfeldt J, Damgaard I, Bryder D, Theilgaard-Monch K, Thoren LA и др. (2008) NMD важен для гемопоэтических стволовых клеток и клеток-предшественников, а также для устранения побочных продуктов запрограммированных перестроек ДНК. Гены Дев 22: 1381–1396. - 4.Brogna S, Wen J (2009) Механизмы нонсенс-опосредованного распада мРНК (NMD). Nat Struct Mol Biol 16: 107–113.
- 5.
Carter MS, Li S, Wilkinson MF (1996) Зависимый от сплайсинга регуляторный механизм, который обнаруживает сигналы трансляции. EMBO J 15: 5965–5975. - 6.
Zhang J, Sun X, Qian Y, Maquat LE (1998) Функция интрона в бессмысленном распаде мРНК бета-глобина: признаки того, что сплайсинг пре-мРНК в ядре может влиять на трансляцию мРНК в цитоплазме.РНК 4: 801–815. - 7.
Терманн Р., Ной-Йилик Дж., Детерс А., Фреде Ю., Вер К. и др. (1998) Бинарная спецификация бессмысленных кодонов путем сплайсинга и цитоплазматической трансляции. EMBO J 17: 3484–3494. - 8.
Le Hir H, Gatfield D, Izaurralde E, Moore MJ (2001) Комплекс экзон-экзонного соединения обеспечивает платформу связывания для факторов, участвующих в экспорте мРНК и нонсенс-опосредованном распаде мРНК. EMBO J 20: 4987–4997. - 9.
Tange TO, Nott A, Moore MJ (2004) Постоянно растущие сложности комплекса соединений экзонов.Curr Opin Cell Biol 16: 279–284. - 10.
Nagy E, Maquat LE (1998) Правило для положения терминирующего кодона в генах, содержащих интрон: когда бессмыслица влияет на количество РНК. Тенденции Biochem Sci 23: 198–199. - 11.
Zhang J, Sun X, Qian Y, LaDuca JP, Maquat LE (1998) По крайней мере, один интрон необходим для нонсенс-опосредованного распада мРНК триозофосфат-изомеразы: возможная связь между ядерным сплайсингом и цитоплазматической трансляцией. Mol Cell Biol 18: 5272–5283. - 12.
Buhler M, Steiner S, Mohn F, Paillusson A, Muhlemann O (2006) EJC-независимая деградация бессмысленной мРНК иммуноглобулина-mu зависит от длины 3 ‘UTR. Nat Struct Mol Biol 13: 462–464. - 13.
Wang J, Gudikote JP, Olivas OR, Wilkinson MF (2002) Гранично-независимый полярный нонсенс-опосредованный распад. EMBO Rep 3: 274–279. - 14.
Cheng J, Belgrader P, Zhou X, Maquat LE (1994) Интроны являются цис-эффекторами опосредованного нонсенс-кодонами снижения численности ядерной мРНК.Mol Cell Biol 14: 6317–6325. - 15.
Matsuda D, Hosoda N, Kim YK, Maquat LE (2007) Отказоустойчивый нонсенс-опосредованный распад мРНК не обнаруживает обнаруживаемой мишени для мРНК, связанной с eIF4E. Nat Struct Mol Biol 14: 974–979. - 16.
Neu-Yilik G, Gehring NH, Thermann R, Frede U, Hentze MW и др. (2001) Сплайсинг и образование 3′-концов в определении нонсенс-опосредованных распадом компетентных мРНП бета-глобина человека. EMBO J 20: 532–540. - 17.
Brocke KS, Neu-Yilik G, Gehring NH, Hentze MW, Kulozik AE (2002) Ген безинтронного рецептора меланокортина 4 человека нечувствителен к NMD.Hum Mol Genet 11: 331–335. - 18.
Maquat LE, Li X (2001) Транскрипты теплового шока млекопитающих p70 и гистона h5, которые происходят из естественно лишенных интронов генов, невосприимчивы к нонсенс-опосредованному распаду. РНК 7: 445–456. - 19.
Chemin G, Tinguely A, Sirac C, Lechouane F, Duchez S и др. (2010) Множественные механизмы наблюдения за РНК взаимодействуют, чтобы уменьшить количество нефункциональных транскриптов каппа Ig. J Immunol 184: 5009–5017. - 20.
Chang YF, Chan WK, Imam JS, Wilkinson MF (2007) Транскрипты альтернативно сплайсированных Т-клеточных рецепторов активируются в ответ на нарушение либо элементов сплайсинга, либо рамки считывания.J Biol Chem 282: 29738–29747. - 21.
Valentine CR (1998) Связь бессмысленных кодонов с пропуском экзонов. Mutat Res 411: 87–117. - 22.
Maquat LE (2002) NASty-эффекты на сплайсинг пре-мРНК фибриллина: это происходит в другом случае ESE, но предложения по выбору сайта сплайсинга, зависящего от трансляции, продолжаются. Гены Дев 16: 1743–1753. - 23.
Ван Дж., Чанг Ю.Ф., Гамильтон Д.И., Уилкинсон М.Ф. (2002) Измененный сплайсинг, связанный с бессмыслицей: зависимый от кадра ответ, отличный от опосредованного нонсенсом распада.Mol Cell 10: 951–957. - 24.
Imam JS, Gudikote JP, Chan WK, Wilkinson MF (2010) Мутации, разрушающие каркас, вызывают накопление пре-мРНК независимо от нарушения каркаса. Nucleic Acids Res 38: 1559–1574. - 25.
Lozano F, Maertzdorf B, Pannell R, Milstein C (1994) Низкие цитоплазматические уровни мРНК генов легкой цепи каппа иммуноглобулина, содержащих нонсенс-кодоны, коррелируют с неэффективным сплайсингом. EMBO J 13: 4617–4622. - 26.
Aoufouchi S, Yelamos J, Milstein C (1996) Нонсенс-мутации ингибируют сплайсинг РНК в бесклеточной системе: распознавание мутантного кодона не зависит от синтеза белка.Ячейка 85: 415–422. - 27.
Картер М.С., Доскоу Дж., Моррис П., Ли С., Нхим Р.П. и др. (1995) Регуляторный механизм, который обнаруживает преждевременные бессмысленные кодоны в транскриптах Т-клеточных рецепторов in vivo, отменяется ингибиторами синтеза белка in vitro. J Biol Chem 270: 28995–29003. - 28.
Glusman G, Rowen L, Lee I, Boysen C, Roach JC и др. (2001) Сравнительная геномика локусов рецепторов Т-клеток человека и мыши. Иммунитет 15: 337–349. - 29.
Brady BL, Steinel NC, Bassing CH (2010) Исключение аллелей антигенных рецепторов: обновление и переоценка.J Immunol 185: 3801–3808. - 30.
Хор Б., Слекман Б. П. (2005) Внутри- и межаллельное упорядочение сборки гена бета-цепи рецептора Т-клеток. Eur J Immunol 35: 964–970. - 31.
Delpy L, Sirac C, Magnoux E, Duchez S, Cogne M (2004) Надзор за РНК подавляет экспрессию нефункциональных аллелей каппа и обнаруживает преждевременное завершение в последнем экзоне каппа. Proc Natl Acad Sci U S A 101: 7375–7380. - 32.
Bassing CH, Alt FW, Hughes MM, D’Auteuil M, Wehrly TD и др.(2000) Сигнальные последовательности рекомбинации ограничивают хромосомную рекомбинацию V (D) J за пределами правила 12/23. Nature 405: 583–586. - 33.
Khor B, Mahowald GK, Khor K, Sleckman BP (2009) Функциональное перекрытие в цис-действующей регуляции рекомбинации V (D) J в локусе TCRbeta. Мол Иммунол 46: 321–326. - 34.
Mahowald GK, Baron JM, Mahowald MA, Kulkarni S, Bredemeyer AL, et al. (2009) Аберрантно разрешенные RAG-опосредованные разрывы ДНК в Atm-дефицитных лимфоцитах нацелены на хромосомные точки разрыва в цис.Proc Natl Acad Sci U S A 106: 18339–18344.
Композиционные характеристики реаранжировок псевдогена периферических TRBV человека
Частота гена
Список данных последовательностей из 4 здоровых образцов и 4 пациентов показан в таблице 1. Последовательности без результатов или Неизвестная или идентичность V-области <85% были отфильтрованы вне. Наконец, мы проанализировали все функциональные (F) перестройки TRBV, перестройки ORF TRBV и перестройки псевдогена (P) TRBV (Таблица 1).Сначала мы исследовали частоты этих трех классов перегруппировок (рис. 1А, Б). Учитывая размер выборки и некоторые экстремальные значения, мы использовали медианное значение для оценки этих данных. На рисунке 1А показано, что частота реаранжировок TRBV P и ORF (медиана 1,0% и 0,7% в здоровой группе; медиана 2,3% и 0,9% в группе заболевания) была намного ниже, чем частота реаранжировок F (медиана 85,6% в группе больных). здоровая группа и медиана 88,7% в группе болезни). В здоровой группе частота псевдогена TRBV21-1 была выше, чем сумма частот других псевдогенов (0.7% против 0,3%), тогда как другие псевдогены не были обнаружены из-за низкой частоты (функциональная подгруппа генов охватывала 45 генов). Это несоответствие увеличилось в группе заболевания из-за распространения TRBV21-1 (1,9% против 0,4%), что также может быть связано с заболеванием. В отличие от реаранжировок P и ORF, очевидно, что реаранжировки F используются по-разному между здоровыми и больными группами (рис. 1A). Рисунок 1B показывает, что перегруппировки F в основном являются продуктивными перегруппировками, в то время как перегруппировки P и ORF характеризуются высокой долей непродуктивных перегруппировок.Взятые вместе, эти результаты предполагают, что перестройки псевдогена TRBV и функциональные перестройки генов могут иметь значительные различия в составе, и побудили нас сравнить их композиционные характеристики.
Таблица 1 Количество последовательностей в различных категориях, идентифицированных у 4 здоровых людей и 4 пациентов. Рисунок 1
Частота гена функциональных (F) генов TRBV, открытых рамок считывания (ORF) TRBV и псевдогенов TRBV (P) в группах здоровых и больных.( A ) Частота функциональных генов TRBV значительно выше, чем частота псевдогенов TRBV и ORF. ( B ) Псевдогены TRBV и ORF демонстрируют непродуктивные перестройки, которые используются в значительной степени, в то время как большинство функциональных генов являются продуктивными. Эти результаты отображаются в виде прямоугольной диаграммы, на которой показаны все точки для обозначения каждого образца. Линия указывает на медианное значение, а две планки ошибок используются для представления минимального и максимального значений.
Использование генов J и D
Локус гена TCRB человека содержит 13 сегментов TRBJ и 2 сегмента TRBD (№ D представляет несколько реаранжировок V-J без сегмента TRBD).Было продемонстрировано, что использование TRBJ неслучайно в продуктивных транскриптах, происходящих из Т-клеток периферической крови 6 . Мы сравнили использование генов J и D для реаранжировок P, ORF и F у здоровых и больных групп (рис. 2A – E). В группах здоровых и больных мы наблюдали значительные различия в общем использовании TRBJ среди P, ORF и F перегруппировок (критерий χ2, 12 степеней свободы, каждый p <0,001), тогда как использование TRBJ1-3, TRBJ1-6 и TRBJ 2-3 были очень похожи (рис.2А, Б). Кроме того, TRBJ2-7 использовался с относительно высокой частотой (каждый процент> 20%) среди реаранжировок P, ORF и F в группах здоровых и больных. Примечательно, что для здоровой группы частота использования TRBJ1-1 в перегруппировках P была значительно ниже, чем в реаранжировках ORF и F (критерий χ2, каждый p <0,01), а реаранжировка P показала более частое использование TRBJ1-5. чем ORF и F перегруппировки (критерий χ2, p <0,05 и p <0,01, соответственно). Кроме того, для группы заболевания частота использования TRBJ1-5 в реаранжировках P была значительно ниже, чем в реаранжировках ORF и F (критерий χ2, каждый p <0.01) (рис. 2А, Б). Мы также обнаружили несколько различий в использовании TRBJ реаранжировок P между здоровыми и больными группами, которые были воплощены в TRBJ1-1, TRBJ1-2, TRBJ1-5, TRBJ2-1 и TRBJ2-6 (рис. 2C). Кроме того, использование TRBD1 и TRBD2 реаранжировок P было относительно равным в группах здоровых и больных (45,3% против 44,4% и 48,9% против 45,9% соответственно), в то время как несоответствие в различиях в использовании TRBD1 или TRBD2 появилось между группами группы здоровых и больных (рис. 2D, E).Хотя было несколько различий между реаранжировками P, ORF и F, было трудно различить P-перегруппировки на основании частот гена J или D.
Рисунок 2
Использование генов TRBJ и TRBD у 4 здоровых людей и 4 пациентов. ( A , B ) Использование генов TRBJ показывает значительные различия между тремя классами перестроек в группах здоровых и больных. Результаты показаны со средним значением в каждой группе.Значения p были определены с помощью критерия χ2 с использованием медианы четырех образцов в каждой группе. Указаны все статистически значимые различия. * p <0,05, ** p <0,01, *** p <0,001. ( C ) Показано сравнение использования TRBJ в псевдогене между здоровыми и больными группами. ( D , E ) Показано использование генов TRBD в трех классах перестроек в группах здоровых и больных. Результаты ( C — E ) нанесены на прямоугольную линейную диаграмму, на которой показаны все точки для обозначения каждого образца в каждой группе.Линия указывает медианное значение, а две планки ошибок представляют минимальное и максимальное значение. Значения p были определены с помощью критерия χ2 с использованием медианы четырех образцов в каждой группе. Указаны все статистически значимые различия. * p <0,05, ** p <0,01, *** p <0,001.
Предполагаемая композиция CDR3 AA перестройки в кадре
Последовательность AA, кодируемая областью соединения V-D-J, очевидно, сильно зависит от используемых последовательностей зародышевой линии TRBD и TRBJ 6 .Принимая во внимание влияние заболевания, мы сравнили области CDR3 между перестройками псевдогена в рамке считывания и функциональными перестройками генов в рамке считывания в группе здоровых людей. Количество уникальных последовательностей CDR3 в каждом образце показано в таблице 2. В группе здоровых мы обнаружили, что средняя длина CDR3 в перестройках псевдогена TRBV (13,5 ± 2,4 остатков АА) была немного больше, чем в функциональных перестройках генов. (12,5 ± 1,8 остатков АА) (рис. 3А). В целом, использование АК в этих уникальных последовательностях CDR3 показало более частое использование аргинина, лизина и цистеина в перестройках псевдогена TRBV (критерий χ2, каждый p <0.001), в то время как больше лейцина было обнаружено при функциональных перестройках генов (критерий χ2, p <0,01) (рис. 3B). Частоты использования других AA были одинаковыми в этих двух группах (рис. 3B). Интересно, что распределение длин CDR3 и общее использование AA перегруппировок псевдогена в рамке считывания между здоровой и больной группами были очень похожими (рис. 3C, D).
Таблица 2 Количество уникальных последовательностей CDR3 от 4 здоровых людей. Фигура 3
длин CDR3 и общее использование аминокислот в областях CDR3 в функциональных перестройках гена TRBV в рамке считывания и перестройках псевдогена TRBV в рамке считывания.( A ) Показано распределение средней длины CDR3 функциональных реаранжировок гена TRBV в рамке считывания и реаранжировок псевдогена TRBV в рамке считывания в здоровой группе. ( B ) Аминокислоты расположены по значениям относительной гидрофобности согласно шкале Кайта-Дулиттла 13 . Общий аминокислотный состав CDR3 значительно отличался между перестройками псевдогена и функциональными перестройками генов в здоровой группе (критерий χ2, p <0,001). ( C , D ) Сравнение среднего распределения длин CDR3 и общего аминокислотного состава CDR3 перегруппировок псевдогена TRBV в рамке считывания между здоровой и больной группами не показывает различий.Результаты показаны как среднее значение в каждой группе. Значения p определяли с использованием критерия χ2 ( B и D ). Указаны все статистически значимые различия. * p <0,05, ** p <0,01, *** p <0,001.
Когда АК были сгруппированы в соответствии с их физическими свойствами, сравнение функциональных генных (выбранных) и псевдогенных (невыбранных) реаранжировок показало, что последовательности псевдогена чаще кодируются для основных (положение 108) и потенциальных остатков компонентов дисульфидного мостика ( позиции 108 и 109) (критерий χ2, каждый p <0.001, рис.4). Уникальная нумерация IMGT используется для идентификации позиции 14,15,16 .
Фигура 4
Частоты аминокислот в десяти конкретных положениях, выбранных для оценки функциональных перестроек гена TRBV в рамке считывания, по сравнению с предполагаемым использованием аминокислот в перестройках псевдогена TRBV в рамке считывания, сгруппированных в соответствии с физическими свойствами. Позиции CDR3 (105–117) показаны в соответствии с уникальной нумерацией IMGT. Результаты нанесены на тепловую карту, которая показывает среднее у 4 здоровых людей всех уникальных продуктивных последовательностей CDR3 разной длины.
Кроме того, мы рассчитали общие частоты AA для 10 позиций в последовательностях CDR3 (рис. S1). Позиции 105, 106, 107, 116 и 117 считались эволюционно сохраненными. Интересно, что значительные различия в положениях 105 и 106 наблюдались между перестройками псевдогена и функциональными перестройками генов. В позиции 105 большее количество псевдогена CDR3 использовало треонин (9,2% против 0,03%), тогда как более частое использование серина наблюдалось в функциональном гене CDR3 (12,2% против 0%).В положении 106 аргинин и серин чаще использовались в перестройках псевдогена TRBV (2,9% против 0,4% и 93,2% против 76,8% соответственно), тогда как триптофан, треонин, аланин и валин использовались реже.
Вставка и делеция нуклеотидов
Разнообразие CDR3 является результатом добавления нуклеотидов «N» в соединениях V → D (N1) и D → J (N2), обрезки экзонуклеаз (3’V обрезано, 5′D обрезано и 5’J обрезано) и добавление палиндромных «P» нуклеотидов (P3’V, P5’D и P5’J).Учитывая функциональность последовательности, мы сравнили вставки и делеции между полными перестройками псевдогена TRBV и функциональными перестройками гена TRBV в здоровой группе (рис. S2). Мы обнаружили, что среднее количество нуклеотидов, добавленных в N1 или N2 при перестройках псевдогена TRBV, было более чем на 1 нуклеотид больше, чем добавленное при функциональных перестройках гена TRBV (ANOVA с поправкой Бонферрони, p <0,001), в то время как существенной разницы в Использование четырех оснований в N1 или N2 наблюдалось между перестройками псевдогена TRBV и функциональными перестройками гена TRBV (рис.S2). Были аналогичные добавления в P-областях (P3'V, P5'D и P5'J) и делеции в 3'V, 5'D и 5'J за счет обрезки экзонуклеаз (фиг. S2). Кроме того, мы удалили фоновый шум последовательности, чтобы проанализировать уникальные последовательности в трех классах перегруппировок. Из-за различных профилей распределения нуклеотидных вставок или делеций в различных функциональных генах TRBV или TRBJ 11 , доля TRBV21-1, обнаруженная в группе псевдогенов, была относительно высокой; поэтому мы наблюдали профили внедрения N TRBV21-1 и no-TRBV21-1 (рис.S3). Примечательно, что подобно функциональным генам, разные псевдогены TRBV обнаруживают разные профили вставки (Fig. S3).
В соответствии с методом классификации вставки и делеции нуклеотидов, описанным Murugan et al . 11 , мы исследовали профили распределения нуклеотидов среди псевдогенов, открытых рамок считывания и функциональных генов (рис. 5A, D).
Рис. 5
Вставки и делеции нуклеотидов в соединениях V-D-J реаранжировок P, ORF и F в здоровой (P-H, ORF-H и F-H) и больной группе (P-D, ORF-D и F-D).( A ) Частотные профили вставок VD различной длины нуклеотидов в трех классах перегруппировок. ( B ) Частотный профиль вставок DJ различной длины нуклеотидов в трех классах перегруппировок. ( C ) Частотный профиль V-делеций разной длины нуклеотидов в трех классах перегруппировок. ( D ) Частотный профиль делеций J разной длины нуклеотидов в трех классах перегруппировок.Результаты показаны как среднее значение всех уникальных непродуктивных последовательностей в каждой группе.
Профиль инсерции VD был сходным для генов P, ORF и F как в группе здоровых, так и в группе больных (фиг. 5A). Профиль вставки DJ перегруппировок F отличался от профилей других групп, что может быть связано с заболеванием (рис. 5B). Примечательно, что профили V-делеции нуклеотидов среди генов P, ORF и F явно различались как в группе здоровых, так и в группе больных (рис. 5C). Кроме того, профили V-делеции одного и того же типа генов были сходными в группах здоровых и больных.Следовательно, характеристики профиля V-делеции нуклеотидов могут отличать репертуар псевдогена от репертуаров ORF и F. Здесь характеристики V-делеционного профиля псевдогенов в основном отражают высокую долю коротких нуклеотидных делеций (1 н.) И высокую частоту нуклеотидных делеций (более 90%).
Балаган представляет … Перестановка кадров и воспоминаний: Японское экспериментальное кино, 1970-е годы — сегодня
Балаган представляет программу редко просматриваемого японского экспериментального кино за последние несколько десятилетий, приглашенным куратором которой является режиссер Томонари Нисикава .Программа ориентирована на короткие произведения с уникальными визуальными эффектами. Некоторые выражают границу между реальностью и иллюзией, в то время как другие просто выражают интересы художников в манипуляциях с изображениями и исследовании границ среды. Фильмы будут представлены в оригинальном формате, при этом большая часть репродукций будет напрямую из Японии.
О кураторе:
Томонари Нисикава (Tomonari Nishikawa) — кинорежиссер и художник-инсталлятор, работающий в форматах 35 мм, 16 мм и Super-8.Его работы демонстрировались на многочисленных фестивалях по всему миру, включая Международный кинофестиваль в Торонто, Международный кинофестиваль в Эдинбурге, кинофестиваль в Анн-Арборе и Берлинале. В качестве куратора он был программистом Международного фестиваля искусства и альтернативного видения Yebisu в Токио. В настоящее время он живет и преподает в Колгейтском университете в северной части штата Нью-Йорк.
Программа:
— Pulse , Keiichi Minegishi, 1982
6 мин, 16 мм
— Cessna , Такаши Накадзима, 1974
20 мин, 16 мм
— Film Display , Shunzo Seo, 1979
5 мин., 16 мм
— Стеклоочиститель , Itaru Kato, 1985
7 мин, 16 мм
— Tokio House , Сумиаки Исида, 1990
7 мин, 16 мм
— Land , Moe Suziki, 2011
8 мин, видео
— Space is the Place , Эрико Сонода, 2011
6 мин, видео
— Кресло-качалка , Шихо Кано, 2000
13 мин, 16 мм
— Shishosetsu , Nobuhiro Kawanaka, 1987
8 мин, 16 мм
Это мероприятие стало возможным благодаря щедрой поддержке Японского фонда, Ассоциации аспирантов колледжа Эмерсон и Университета Колгейт.
Тяжелые цепи иммуноглобулина — обзор
5 GC Response
Рекомбинация переключения класса тяжелой цепи иммуноглобулина происходит в GC B-клетках с трансформацией IgM-продуцирующих B-клеток в IgG-продуцирующие B-клетки, изменяя эффекторную функцию антитела молекула. Это имеет отношение к аутоиммунитету, поскольку антитела IgM, которые связывают апоптотический мусор и другие эндогенные лиганды для TLR, образуют иммунные комплексы, которые опосредуют иммуносупрессивный эффект, частично за счет вовлечения C1q, который затем связывает LAIR-1, ингибирующий рецептор на моноцитах и DC (Gronwall). , Vas, & Silverman, 2012; Peng, Kowalewski, Kim, & Elkon, 2005; Roos et al., 2004; Сын, Сантьяго-Шварц, Аль-Абед и Даймонд, 2012 г.). Напротив, антитела IgG, отличающиеся от своих аналогов IgM только изотипом тяжелой цепи, образуют иммунные комплексы, которые могут взаимодействовать с активирующими рецепторами Fc на клетках миелоидного клона и тем самым создавать провоспалительную среду, в которой аутоантигены могут быть представлены иммуногенным образом. В то время как В-клетки экспрессируют только ингибирующий рецептор Fc, FcγRllB, его участие иммунными комплексами IgG, но не иммунными комплексами IgM, может помочь транспортировать лиганды TLR в В-клетки для запуска программ выживания и активации в клетке, которая в противном случае могла бы подвергнуться толерантности или оставаться в состоянии покоя. (Green & Marshak-Rothstein, 2011; Herlands, Christensen, Sweet, Hershberg, & Shlomchik, 2008; Pasare & Medzhitov, 2005).Таким образом, аутоантитела IgG могут создавать петлю амплификации, когда они созданы, что увеличивает аутореактивность. В то время как рекомбинация переключения класса тяжелой цепи может быть индуцирована вне GC в отсутствие взаимодействия родственных Т-клеток, например, с помощью BAFF, IFNγ, IFN типа 1 или IL-21, рекомбинация переключения тяжелой цепи является одним из ключевых аспектов ответ GC.
Точечные мутации в генах вариабельной области иммуноглобулина (тяжелая цепь чаще, чем легкая цепь) в ответе GC приводят ко второй волне диверсификации репертуара антител с последующим отбором В-клеток с высоким сродством к вызывающему антигену.Именно те В-клетки, которые позитивно отбираются в ГК, могут стать долгоживущими плазматическими клетками в костном мозге или В-клетками памяти (Berek, Berger, & Apel, 1991; обзор в Chan & Brink, 2012). Изменения в экспрессии белка внутри GC B-клетки способствуют образованию B-клеток памяти и долгоживущих плазматических клеток, которые экспрессируют высокоаффинные антитела с переключенным классом. AID имеет решающее значение как для рекомбинации с переключением классов, так и для соматической гипермутации, ключевых процессов ответа GC (Hase et al., 2008; Мурамацу и др., 2000; Захин и др., 2009; Захин и Мартин, 2011). Генерация GC-созревших В-клеток имеет решающее значение для защитного иммунитета. Пациенты без AID имеют интактные низкоаффинные IgM-ответы, но неспособны развивать B-клеточную память и высокоаффинные антитела с переключением классов тяжелых цепей (Revy et al., 2000). В то время как AID может экспрессироваться в В-клетках, не являющихся GC, а рекомбинация с переключением класса тяжелых цепей и соматическая гипермутация действительно происходят вне среды GC, уровни экспрессии и функции AID являются самыми высокими в GC (Zaheen & Martin, 2011).
Роль СПИДа в возникновении аутоиммунитета сложна. Имеются веские доказательства того, что высокие уровни AID связаны с аутоиммунитетом как у мышей, так и у пациентов с RA, у которых циркулирующие B-клетки экспрессируют высокие уровни AID (Hsu et al., 2007; Jiang et al., 2007; Xu et al., 2009). И наоборот, дефицит СПИДа у мышей и людей часто связан с аутоиммунитетом; происходит ли это у мышей, зависит от генетического фона (Hase et al., 2008; Quartier et al., 2004). GCs образуются у AID-дефицитных мышей, но GC B-клетки у этих мышей неспособны подвергаться апоптозу (Zaheen et al., 2009). Следовательно, аутоиммунитет мышей с дефицитом AID приписывают неспособности апоптоз-зависимого отбора GC B-клеток.
Экспрессия BCR снижена в GC B-клетках, а экспрессия Fas повышена (Bras, Martinez, & Baixeras, 1997; Koncz & Hueber, 2012). Эта совокупность изменений важна для созревания аффинности ответа антител. BCR, по-видимому, функционирует больше, чтобы облегчить адгезию между B-клетками и другими клетками, чтобы увеличить выживаемость и пролиферацию клеток, а не инициировать передачу сигналов BCR (Khalil, Cambier, & Shlomchik, 2012).Фактически, BCR в GC B клетках совместно локализуется с повышенным содержанием фосфатаз SHP-1 и SHIP-1 (Khalil et al., 2012). Активация фосфатаз при вовлечении BCR предотвращает сильную передачу сигналов BCR. Кроме того, снижение экспрессии BCR в GC B-клетках усиливает конкуренцию за антиген. Высокий уровень экспрессии Fas гарантирует, что В-клетки, которые не испытывают взаимодействия с BCR, разрушаются посредством апоптотического пути (обзор Peperzak, Vikstrom, & Tarlinton, 2012).Ясно, что множественные подкрепляющие сигналы управляют выживанием и пролиферацией антиген-специфических В-клеток и необходимы на протяжении всего времени, пока В-клетка находится в среде GC.
По мере накопления точечных мутаций в сегментах гена иммуноглобулина V образуются антитела, которые имеют более высокое сродство к вызывающему антигену. Те В-клетки с достаточной начальной стимуляцией как антигеном, так и Т , FH и FDC пролиферируют и подвергаются соматической гипермутации. В-клетки, подвергшиеся соматической мутации, конкурируют друг с другом за помощь антигена и родственных Т-клеток (Batista & Neuberger, 2000).Таким образом происходит положительный отбор высокоаффинных антител (обзор в Zotos & Tarlinton, 2012). Те В-клетки с наивысшим сродством к антигену, скорее всего, станут плазматическими клетками, а не клетками памяти (Smith, Light, Nossal, & Tarlinton, 1997; Smith et al., 2000). Было высказано предположение, что сильное взаимодействие BCR приводит к деградации Bcl-6 и, как следствие, дерепрессии Blimp-1, что приводит к дифференцировке плазматических клеток (Shapiro-Shelef et al., 2003; Shapiro-Shelef, Lin, Savitsky, Liao, И Calame, 2005).Однако антитела с меньшим сродством к элисирующему антигену или без него, а также антитела, которые приобретают аутореактивность, которая может или не может быть перекрестно реактивной с элисирующим антигеном, также образуются во время ответа GC.
То, что соматическая мутация может приводить к приобретению аутоспецифичности, было впервые продемонстрировано нами в исследованиях линии клеток миеломы мышей. Клеточная линия S107 вырабатывает канонические антитела к фосфорилхолину, доминирующему эпитопу на полисахариде клеточной стенки пневмококка, который защищает мышей от летальной пневмококковой инфекции.Простая замена основания в вариабельной области тяжелой цепи антитела приводит к антителу с заметно сниженным связыванием с фосфорилхолином, но с новой реактивностью по отношению к ДНК (Diamond & Scharff, 1984). Это наблюдение впервые позволило предположить, что аутореактивность может возникать из-за соматической мутации, в отличие от ранее существовавшей точки зрения, согласно которой соматическая мутация является механизмом устранения аутореактивности.
За этим наблюдением последовало исследований in vivo и на мышах, подтверждающих, что В-клетки могут приобретать аутореактивность в процессе соматической мутации.Мы предположили, что В-клетки, приобретающие аутореактивность в ответе на GC, могут регулярно подвергаться апоптозу, чтобы предотвратить проявление патогенной аутореактивности. Поэтому мы выполнили слияние B-клеток селезенки после иммунизации мышей BALB / c фосфорилхолином, связанным с белком-носителем, с использованием партнера слияния, сконструированного для сверхэкспрессии Bcl-2 и для предотвращения апоптоза гибридом, образованных с помощью B-клеток, запускаемых in vivo подвергнуться апоптозу (Ray, Putterman, & Diamond, 1996).Примерно 40% гибридом, связывающих фосфорилхолин, проявляют перекрестную реактивность с ДНК. Это исследование продемонстрировало высокую частоту аутореактивности в ответе GC. Мансер и его коллеги аналогичным образом продемонстрировали анти-ДНК-реактивность, возникающую внутри ГК в ответ на гаптен фениларсонат (Alabyev, Rahman, & Manser, 2007).
Эти наблюдения привели к исследованиям аутоантител, проведенным пациентами с аутоиммунным заболеванием, чтобы увидеть, закодирована ли их аутореактивность в последовательности зародышевой линии или была приобретена в результате соматической мутации.Мы и другие продемонстрировали, что антитела против ДНК демонстрируют высокую частоту соматических мутаций и что мутации ответственны за приобретение автоспецифичности (Detanico et al., 2012; Jacobi, Hansen, Burmester, Dorner, & Lipsky, 2000; Manheimer- Lory, Zandman-Goddard, Davidson, Aranow, & Diamond, 1997; Mietzner et al., 2008; Schroeder, Herrmann, & Winkler, 2013). Действительно, недавнее исследование В-клеток памяти в крови здоровых людей показывает, что примерно 15% являются аутореактивными В-клетками, что больше, чем в репертуаре наивных В-клеток (Mietzner et al., 2008). Положительный отбор аутореактивного подмножества, по-видимому, не объясняет увеличения аутореактивности между наивным компартментом и компартментом памяти, поскольку по крайней мере половина аутореактивных B-клеток в памяти приобретает свою реактивность в процессе соматической гипермутации внутри GC. Таким образом, похоже, что аутореактивность обычно генерируется GC-зависимой волной диверсификации BCR.
Другое недавнее исследование показывает более низкую частоту аутореактивных В-клеток в пуле плазматических клеток костного мозга, чем пул клеток памяти крови (Scheid et al., 2011). Хотя изученные субпопуляции B-клеток были получены от разных пациентов, одним из выводов данных является то, что может существовать контрольная точка толерантности, прежде чем они станут долгоживущими плазматическими клетками. Альтернативно, может иметь место специфическая потеря аутореактивных В-клеток в популяции плазматических клеток; возможно, что включение рецептора Fc иммунными комплексами, содержащими аутоантиген, приводит к избирательной гибели аутореактивных плазматических клеток (Fukuyama, Nimmerjahn, & Ravetch, 2005; Tzeng, Bolland, Inabe, Kurosaki, & Pierce, 2005).
Сортировка кадра данных в R (4 примера)
В этой статье объясняется, как отсортировать фрейм данных на языке программирования R.
Я покажу вам четыре различных примера для упорядочивания кадров данных. Точнее, учебник будет содержать следующее содержание:
Приступим…
Создание примера данных
Мы собираемся использовать следующий набор данных для примеров этого учебника R:
данные <- данные.frame (x1 = 1: 5, # Создать пример данных
x2 = c («A», «D», «C», «A», «d»))
data # Распечатать пример данных |
data <- data.frame (x1 = 1: 5, # Создать пример данных x2 = c («A», «D», «C», «A», «d»)) data # Пример печати data
Таблица 1: Пример кадра данных.
Таблица 1 показывает, как выглядит наш пример фрейма данных. Наши данные содержат два столбца (т.е. x1 и x2) и пять строк.
Таблица 2 показывает, как следует переставить данные нашего примера, если мы хотим упорядочить их в соответствии со столбцом x2. Вторую строку нужно переместить вниз, а четвертая строка должна стать второй строкой:
Таблица 2: Как следует упорядочить фрейм данных.
Давайте сделаем эту перестановку в R…
Пример 1: Сортировка кадра данных с базой R (функция порядка)
Установка Base R уже предоставляет хорошее решение для упорядочивания наших данных.Нам просто нужно применить функцию порядка к вектору-столбцу, в соответствии с которым мы хотим отсортировать наши данные (например, x2). Взгляните на следующий код R:
data [order (data $ x2),] # Данные заказа с Base R |
data [order (data $ x2),] # Данные заказа с Base R
Таблица 3: Заказанный фрейм данных.
Как видно из таблицы 3, наши данные идеально упорядочены по отношению ко второму столбцу.
Однако, в зависимости от ваших личных предпочтений, вы можете предпочесть другое решение для сортировки ваших данных. Поэтому ниже я собираюсь показать вам некоторые программные альтернативы для упорядочивания фреймов данных ...
Пример 2: Сортировка кадра данных с помощью пакета dplyr (функция упорядочивания)
Пакет dplyr - очень мощный пакет для управления фреймами данных, и пакет также предоставляет функции для упорядочивания матрицы данных.Давайте установим и загрузим пакет в R:
install.packages ("dplyr") # Установить пакет dplyr R
library ("dplyr") # Загрузить пакет dplyr R |
install.packages ("dplyr") # Установить пакет dplyr R
library ("dplyr") # Загрузить пакет dplyr R
Теперь мы можем использовать функцию аранжировки пакета dplyr следующим образом:
организовать (данные, x2) # Данные заказа с dplyr |
организовать (данные, x2) # Данные заказа с dplyr
Посмотрите на вывод вашей консоли RStudio: он покажет упорядоченный фрейм данных так же, как мы его отсортировали в примере 1.
Пример 3: Сортировка кадра данных с пакетом data.table (функция setorder)
Другой популярный пакет для обработки наборов данных - пакет data.table. Во-первых, нам нужно установить и загрузить дополнительный пакет R:
install.packages ("data.table") # Установить пакет R data.table
library ("data.table") # Загрузить data.table R package |
install.packages ("data.table ") # Установить пакет data.table R
library ("data.table") # Загрузить data.table R package
В отличие от предыдущих кодов R, пакет data.table автоматически изменяет исходный фрейм данных. Давайте продублируем данные нашего примера, чтобы сохранить исходную версию данных.
data_ordered <- data # Репликация данных примера |
data_ordered <- data # Репликация данных примера
Теперь мы можем отсортировать и распечатать этот фрейм данных с помощью функции setorder следующим образом:
setorder (data_ordered, x2) # Данные заказа с данными.Таблица data_ordered # Распечатать упорядоченные данные |
setorder (data_ordered, x2) # Данные заказа с таблицей data.table
data_ordered # Распечатать упорядоченные данные
Тот же результат, что и раньше - Выглядит хорошо!
Пример 4: Сортировка кадра данных в порядке убывания
До сих пор мы упорядочивали наши данные только в порядке возрастания (то есть от меньших значений к высоким).Однако мы также можем отсортировать фреймы данных в порядке убывания:
data [порядок (данные $ x2, уменьшение = ИСТИНА),] # Упорядочить данные в порядке убывания |
данные [порядок (данные $ x2, уменьшение = ИСТИНА),] # Упорядочить данные в порядке убывания
Как вы можете видеть на основе вывода вашей консоли RStudio, данные нашего примера были упорядочены в алфавитном порядке от старших до младших букв.
Видео и другие ресурсы
Я опубликовал примеры этого урока в видео на YouTube-канале Statistics Globe.Вы можете посмотреть видео ниже:
Пожалуйста, примите файлы cookie YouTube для воспроизведения этого видео. Согласившись, вы получите доступ к контенту YouTube, услуги, предоставляемой третьей стороной.
Политика конфиденциальности YouTube
Если вы примете это уведомление, ваш выбор будет сохранен, и страница обновится.
Принять контент YouTube
Кроме того, вы можете ознакомиться с некоторыми из соответствующих руководств по R на моей домашней странице.Вы можете найти подборку руководств ниже:
Я надеюсь, что это руководство научило вас сортировать строки фрейма данных по значениям столбцов в языке программирования R. Если у вас возникнут дополнительные вопросы, дайте мне знать в комментариях.
/ * Добавьте свои собственные переопределения стиля формы MailChimp в таблицу стилей вашего сайта или в этот блок стилей.
Мы рекомендуем переместить этот блок и предыдущую ссылку CSS в HEAD вашего HTML-файла. * /
]]>
Изменение порядка столбцов DataFrame в Pandas: Недурно стандартным решениям | Адам Росс Нельсон
Фото: https: // www.flickr.com/photos/takomabibelot/
Решение, упрощающее процесс изменения порядка столбцов DataFrame.
Обновление
, февраль 2021 г., см. ниже руководство, доступное на YouTube.
Десятки сообщений в блогах, веток stackoverflow.com, статей quora.com или других ресурсов демонстрируют аналогичные стандартные методы перемещения столбцов (изменения их порядка) в Pandas DataFrame.
В этой статье сначала приводится пример данных. Следуя данным примера, в статье представлены два хороших , но не идеальных стандартных решений .В этой статье обсуждаются ограничения этих хороших, но не идеальных стандартных решений. Наконец, есть пересмотренное решение, которое позволяет избежать некоторых ограничений, присущих стандартным решениям.
import pandas as pd # Запишите пример данных
df = pd.DataFrame ({'names': ['Al', 'Myke', 'Lovie', 'Dash'],
'Grade': [6,7 , 6,6],
'Учитель': ['Ogna', 'Hayes', 'Stoltz', 'Edens'],
'Оценка': ['A +', 'A -', 'B', 'C »],
« Комната »: [« Зеленый »,« Красный »,« Синий »,« Желтый »],
« Хобби »: [« Монеты »,« Марки »,« Монеты »,« Карты »]})
Пример DataFrame
Явный список столбцов в новом желаемом порядке
Решение, которое чаще всего меняет порядок размещаемых столбцов, состоит в том, чтобы установить DataFrame равным самому себе, но с явно названными столбцами в новом желаемом порядке:
# Reset DataFrame with columns in желаемый порядок
df = df [['name', 'Score', 'Room', 'Grade', 'Teacher', 'Hobby']]
Или просто оценить фрейм данных со столбцами, явно названными в новом желаемом order:
# Оценить DataFrame со столбцами в желаемом порядке
df [['name', 'Score', 'Room', 'Grade', 'Teacher', 'Hobby']]
Это стандартное решение работает хорошо, если там десятки или сотни столбцов.Допустим, у вас есть DataFrame с десятками или сотнями столбцов, и вы хотите переместить только один столбец. Будет неэффективно явно указывать все столбцы для перемещения одного столбца. Перечислить все столбцы также проблематично, поскольку изменения других аспектов связанной базы кода могут вызвать ошибки. Добавление, удаление или изменение имен других столбцов нарушит предыдущие реализации стандартного решения.
Использование удобных методов Pandas
Еще одно стандартное решение - подключить pd.drop () и pd.insert () . Основным ограничением этой опции является то, что она работает только при перемещении одного столбца за раз в определенное место.
mid = df ['Hobby']
df.drop (labels = ['Hobby'], axis = 1, inplace = True)
df.insert (1, 'Hobby', mid)
df
Hobby перемещено из индексировать позицию 5 до позиции индекса 1.
Все еще не идеально, но потенциально лучший вариант для многих случаев использования.
Что делать, если у вас много столбцов, они не расположены рядом друг с другом, и вы хотите переместить их все в новое место после другого определенного столбца? Возможно, вы также захотите одновременно изменить порядок столбцов.Вот решение, которое выполняет поставленную задачу.
def movecol (df, cols_to_move = [], ref_col = '', place = 'After'):cols = df.columns.tolist ()
if place == 'After':
seg1 = cols [: list (cols) .index (ref_col) + 1]
seg2 = cols_to_move
if place == 'Before':
seg1 = cols [: list (cols) .index (ref_col)]
seg2 = cols_to_move + [ref_col]seg1 = [i для i в сегменте 1, если я не в сегменте 2]
сегмент 3 = [i для i в столбцах, если я не в сегменте 1 + сегмент 2]return (df [сегмент 1 + сегмент 2 + сегмент 3])
Для реализации:
df = movecol (df,
cols_to_move = ['Score', 'Grade'],
ref_col = 'Room',
place = 'After')
df
Столбцы Score и Grade изменили свой порядок и переместились в индекс положение после комнаты.
Вообще говоря, управлять данными сложно. После объединения нескольких источников данных удобно иметь метод, который может быстро упорядочить и изменить порядок столбцов.
Это обычная любезность для будущих ученых, которые рассмотрят вашу работу. Если мы разместим наиболее важные столбцы слева, другие будут тратить меньше времени на поиск информации, необходимой для понимания нашей работы. Этот дополнительный шаг также может быть полезен для нас, когда мы знаем, что вернемся к нашей работе для дальнейшего использования.
Если вам нравится то, что я говорю, вы можете найти больше на adamrossnelson.medium.com.
Спасибо за чтение. Присылайте мне свои мысли и идеи. Вы можете написать, просто чтобы поздороваться. И если вам действительно нужно рассказать мне, как я ошибся, я с нетерпением жду встречи в ближайшее время. Twitter: @adamrossnelson | LinkedIn: Адам Росс Нельсон | Facebook: Адам Росс Нельсон.
Изменение порядка столбцов, Учебник на YouTube. Добавлено в феврале 2021 года.
Специальная теория относительности - Перестановка формулы сокращения длины
Важно помнить, что $ S $ может определить длину движущегося стержня только тогда, когда он одновременно измеряет две конечные точки стержня в своей системе отсчета: вот почему мы берем $ t_1 = t_2 $ .2}) \ tag {1} $$
В каждой точке $ x $ кадра $ S $ в каждый момент времени присутствуют два тактовых генератора: один находится в состоянии покоя во время чтения $ S $ $ t $, а другой движется вместе с $ S '$ (прикреплен к $ S '$) время чтения $ t' $, которое в этот момент прошло через $ x $ (синхронные часы меняются от одного момента к другому). Человек, покоящийся в кадре $ S $ $ \ textit {наблюдает} $ 1 , все часы, которые находятся в покое в $ S $, должны быть синхронизированы, а $ \ textit {наблюдать} $ s, чтобы все движущиеся часы были вне sync (Относительность одновременности).
1 Наблюдение - это здесь технический термин: то, что человек $ \ textit {видит} $, не является тем, что он / она $ \ textit {наблюдает} $. При наблюдении учитывается поправка на время прохождения света.
$$ \ underline {\ textbf {Обновленный ответ}} $$
Я думаю, что ваше замешательство, скорее всего, связано с непониманием того, каковы на самом деле величины, связанные с преобразованием Лоренца. Это будет долгое объяснение. Так что держитесь крепче!
$$ \ underline {\ text {(Общий) инструктивный способ визуализации инерциальных кадров}} $$
Рассмотрим для наших целей движение только в направлении $ x $.Вы отдыхаете в кадре $ S $. Представьте себе бесконечно длинную линейку, прикрепленную к раме, с нулевой отметкой, расположенной в вашем местоположении (выбранной вами исходной точке). Кроме того, представьте себе мини-роботов (пожалуйста, продолжайте со мной) с синхронизированными часами, размещенными во всех точках линейки. У этих роботов есть камеры, которые записывают все, что происходит в непосредственной близости от них. Для нас они выглядят как настольные часы, прикрепленные к линейке (очень удобно для наглядности) [см. Рисунок ниже]. Следовательно, когда рамка движется, линейка движется вместе с часами, прикрепленными к линейке.
Что я имею в виду под синхронизированными часами, спросите вы? Выберите любого бота и попросите его кричать вам обоим, расстояние ($ x $) (глядя на показания на стационарной линейке) и показания его часов ($ t $). Допустим, вы получаете это сообщение, когда на ваших часах отображается значение $ (T) $. Вы обнаружите, что величина $ t + ({x \ over c}) $ соответствует $ T $ (здесь $ c $ - это скорость звука в вашем кадре). Если это условие верно для любого бота (и, как следствие, для каждого бота), то часы на ботах синхронизируются относительно вашего кадра.
Вместо того, чтобы писать это пространно, я мог бы просто сказать, что вы $ \ textit {наблюдаете} $, чтобы все часы бота считывали одно и то же время. То, что вы видите, - это не то, что вы $ \ textit {наблюдаете} $, поскольку $ \ textit {Наблюдение} $ учитывает корректировку времени прохождения сигналов. Помните об этой визуализации, чтобы следующие части имели смысл.
$$ \ underline {\ text {Преобразование Лоренца}} $$
Ваш фрейм равен $ S $, а фрейм, движущийся вправо со скоростью $ v $, равен $ S '$.Я назову линейку, прикрепленную к фрейму $ S $, линейку $ S $ и мини-ботов, прикрепленную к линейке $ S $, как $ S $ -боты (и аналогично для $ S '$). Допустим, $ S $ -бот, расположенный в $ x $, записывает показания соседнего движущегося $ S '$ - позиции и времени бота (т.е. $ (x', t ') $), когда его собственные часы показывают $ t $. [См. Рисунок ниже]. Точно так же вы всегда можете представить, что соседний $ S '$ - бот делает то же самое. Преобразование Лоренца просто связывает эти две пары величин: $ (x, t) $ и $ (x ', t') $.
На рисунке ниже показано, что наблюдает покоящийся в кадре $ S $ $ \ textit {} $ человек.
Чтобы ответить на ваш вопрос в комментариях: Почему $ t_1 '\ neq t_2' $?
Посмотрите на рисунок выше и представьте, что концы подвижного стержня расположены в точках $ O '$ и $ x' $. Вы видите, что $ S '$ - часы, расположенные в $ O' $ и $ x '$, показывают разное время? Это то, что я имел в виду, когда сказал, что они не синхронизированы. Часы $ S $ ниже показывают одинаковое время во всех точках линейки $ S $.
Часы $ S '$ синхронизируются в кадре $ S' $, но не синхронизируются в кадре $ S $.Когда человек в состоянии покоя в кадре $ S $ пытается одновременно измерить конечные точки стержня ($ t_1 = t_2 $), он заметит, что $ S '$ - боты, которые находятся рядом с конечными точками стержня, будут читать разные раз ($ t_1 '\ neq t_2' $).
Прошу прощения, если я только запутал вас дальше. Я знаю, что должен быть лучший способ объяснить это, но я имею в виду эту картину, которая лично работает на меня и не дает мне делать ошибок.
Дайте мне знать в комментариях, если у вас есть вопросы / вы найдете что-то, что не имеет смысла в моем объяснении.
.